受體酪氨酸激酶
受體酪氨酸激酶
受體酪氨酸激酶(receptor protein tyrosine kinase,RPTKs)
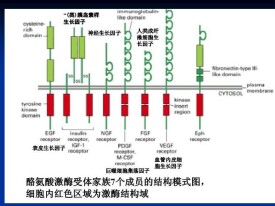
RPTKs是最大的一類酶聯受體,它既是受體,又是酶,能夠同配體結合,並將靶蛋白的酪氨酸殘基磷酸化。所有的RPTKs都是由三個部分組成的:含有配體結合位點的細胞外結構域、單次跨膜的疏水α螺旋區、含有酪氨酸蛋白激酶(PTK)活性的細胞內結構域。
受體酪氨酸激酶(RPTK)是許多多肽生長因子,細胞因子和激素的高親和性細胞表面受體。在人類基因組中鑒定的90種獨特的酪氨酸激酶基因中,有58種編碼受體酪氨酸激酶蛋白。受體酪氨酸激酶不僅被證明是正常細胞過程的關鍵調節因子,而且還在許多類型的癌症的發展和惡化中起關鍵作用。受體酪氨酸激酶的突變會激活一系列信號級聯反應,這些級聯反應對蛋白質表達有很多影響。受體酪氨酸激酶是酪氨酸蛋白激酶家族的一部分,其中包括含有跨膜結構域的受體酪氨酸激酶蛋白,以及不具有跨膜結構域的非受體酪氨酸激酶。
RPTKs的主要類型
目前已知約有50多種,可分為以下20類:
I(EGF受體家族,又稱ErbB受體家族):EGFR, ERBB2, ERBB3, ERBB4
II(胰島素受體家族,Insulin receptor family):INSR,IGFR
III(血小板衍生生長因子受體家族,PDGF receptor family):PDGFRα, PDGFRβ, M-CSFR, KIT, FLT3L
IV(VEGF receptorsfamily):VEGFR1, VEGFR2, VEGFR3
V(成纖維細胞生長因子受體家族,FGF receptor family):FGFR1, FGFR2, FGFR3, FGFR4
VI(CCK receptorfamily):CCK4
VII(神經生長因子受體家族,NGF receptor family):TRKA, TRKB, TRKC
VIII(HGF receptorfamily):MET, RON
IX(Eph receptorfamily):EPHA1 to 6, EPHB1 to 6
X(AXL receptorfamily):AXL, MER, TYRO3
XI(TIE receptorfamily):TIE,TEK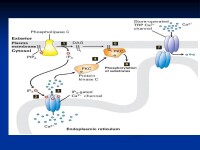
各類受體酪氨酸激酶
XII(RYK receptorfamily):RYK
XIII(DDR receptorfamily):DDR1, DDR2
XIV(RET receptorfamily):RET
XV(ROS receptor family):ROS
XVI(LTK receptorfamily):LTK,ALK
XVII(受體酪氨酸激酶樣孤兒受體家族,ROR receptor family):ROR1, ROR2
XVIII(MuSK receptorfamily):MUSK
XIX(LMR receptor):AATYK1, AATYK2, AATYK3
XX(Undetermined):RTK106
受體酪氨酸激酶在沒有同信號分子結合時是以單體存在的,並且沒有活性;一旦有信號分子與受體的細胞外結構域結合,兩個單體受體分子在膜上形成二聚體,兩個受體的細胞內結構域的尾部相互接觸,激活它們的蛋白激酶的功能,結果使尾部的酪氨酸殘基磷酸化。磷酸化導致受體細胞內結構域的尾部裝配成一個信號複合物(signalingcomplex)。剛剛磷酸化的酪氨酸部位立即成為細胞內信號蛋白(signaling protein)的結合位點,可能有10~20種不同的細胞內信號蛋白同受體尾部磷酸化部位結合后被激活。信號複合物通過幾種不同的信號轉導途徑,擴大信息,激活細胞內一系列的生化反應;或者將不同的信息綜合起來引起細胞的綜合性應答(如細胞增殖)。
通過多種方式,細胞外配體結合通常會引起或穩定受體二聚化。這使得每個受體單體的細胞質部分中的酪氨酸被其伴侶受體反式磷酸化,從而通過質膜傳播信號。活化受體內特定酪氨酸殘基的磷酸化為含有SH2結構域和磷酸酪氨酸結合(PTB)結構域的蛋白提供了結合位點。含有這些結構域的蛋白質包括Src和磷脂酶Cγ。與受體結合的這兩種蛋白質的磷酸化和活化引發了信號轉導通路。與活化受體相互作用的其他蛋白質作為銜接蛋白,並且自身沒有內在酶活性。這些銜接蛋白將RTK活化與下游信號轉導通路聯繫起來,例如MAP激酶信號級聯反應。有一個重要的信號轉導通路包含酪氨酸激酶受體c-met,c-met是在肌肉發生過程中,遷移中的成肌細胞的存活和增殖所必需的。缺乏c-met會破壞繼發性肌細胞生成,並且如同LBX1一樣,可以防止肢體肌肉組織的形成。FGF(成纖維細胞生長因子)與其RTK受體的這種局部作用被歸類為旁分泌信號傳導。由於RTK受體磷酸化多個酪氨酸殘基,它們可以激活多種信號轉導通路。
受體酪氨酸激酶(RTK)途徑受各種正反饋迴路的嚴格調節。因為RTK協調多種細胞功能,例如細胞增殖和分化,所以必須對它們進行調節以防止細胞功能發生嚴重異常,例如癌症和纖維化。
蛋白質酪氨酸磷酸酶(PTP)是一組具有磷酸酪氨酸特異性磷酸水解酶活性的催化結構域的酶。PTP能夠以正向和負向改變受體酪氨酸激酶的活性。PTP可以使RTK上活化的磷酸化酪氨酸殘基去磷酸化,這實際上導致信號終止。有關PTP1B(一種廣為人知的參與細胞周期和細胞因子受體信號調節的PTP)的研究表明,它可以使表皮生長因子受體和胰島素受體去磷酸化。另一方面,一些PTP是在細胞信號增殖中起積極作用的細胞表面受體。細胞表面糖蛋白Cd45在抗原刺激的特異性磷酸酪氨酸的去磷酸化中發揮關鍵作用,而這抑制Src途徑。
赫斯達汀(Herstatin)是ErbB家族的自抑製劑,它與RTK結合併阻斷受體二聚化和酪氨酸磷酸化。用赫斯達汀轉染CHO細胞響應於EGF會導致受體寡聚化、克隆生長和受體酪氨酸磷酸化的減少。
活化的RTK可以進行內吞作用,導致受體數量下調,最終導致信號級聯減弱。其分子機制涉及通過網格蛋白介導的內吞作用吞噬RTK,使其在細胞內降解。
基本信息
- 中文名
- 受體酪氨酸激酶
- 外文名
- receptor tyrosine kinase, RTKs
- 別名
- 抑制酪氨酸激酶活性的化合物
- 警示術語
- 主要通過抑制細胞信號轉導而抑制腫瘤細胞的生長和增殖,促進細胞凋亡
- 類別
- 最大的一類酶聯受體
- 類型
- 表皮生長因子