三域系統
三域系統
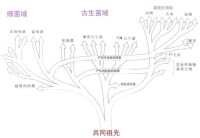
三域系統是由卡爾·烏斯(Carl Woese)在1977年提出的細胞生命形式分類,將原核生物分成了兩大類,起初稱為真細菌(Eubacteria)和古細菌(Archaebacteria)。Woese依據16S rRNA序列上的差別,認為這兩類生物和真核生物一起從一個具有原始遺傳機制的共同祖先分別演化而來,因此將三者各置為一個“域”(Domain),作為比“界”高一級的分類系統,並分別命名為細菌域(Bacteria)、古菌域(Archaea)和真核域(Eukarya)。
然而,rRNA樹有可能將演化太快的類群放錯位置(比如微孢子蟲)。有人認為生物的根應該在真細菌之內,很多真細菌的類群在古細菌之前分支出來,很晚古細菌和真核生物才彼此分開。
三域系統
然而﹐RNA樹有可能將進化太快的類群放錯位置(比如微孢子蟲)。有人認為生物的根應該在真細菌之內﹐很多真細菌的類群在古細菌之前分支出來﹐很晚古細菌和真核生物才彼此分開。
| 三域特徵比較 | ||||
|---|---|---|---|---|
| 特徵 | 細菌(Bacteria) | 古菌(Archaea) | 真核生物(Eukarya) | |
| 形態和遺傳 | 原核細胞結構 | 是 | 是 | 否 |
| 共價閉合環狀DNA | 是 | 是 | 否 | |
| 組蛋白 | 無 | 有 | 有 | |
| 被膜包被的核 | 無 | 無 | 有 | |
| 細胞壁 | 含細胞酸 | 不含細胞酸 | 不含細胞酸 | |
| 膜脂質 | 酯鍵連接 | 醚鍵連接 | 酯鍵連接 | |
| 核糖體大小 | 70S | 70S | 80S | |
| 起始tRNA | 甲醯蛋氨酸 | 蛋氨酸 | 蛋氨酸 | |
| 多數基因中有內含子 | 否 | 否 | 是 | |
| 操縱子 | 有 | 有 | 無 | |
| mRNA加帽和聚A尾 | 無 | 無 | 有 | |
| 質粒 | 有 | 有 | 稀少 | |
| 核糖體對白喉毒素敏感 | 否 | 是 | 是 | |
| RNA聚合酶 | 1個(4亞基) | 幾個(各8~12亞基) | 3個(各12~14亞基) | |
| 需要轉錄因子 | 否 | 是 | 是 | |
| 啟動子結構 | -10和-35序列 | TATA框 | TATA框 | |
| 生理 | 產甲烷作用 | 無 | 有 | 無 |
| 硝化 | 有 | 無 | 無 | |
| 脫氮 | 有 | 有 | 無 | |
| 固氮 | 有 | 有 | 無 | |
| 基於葉綠素的光合作用 | 有 | 無 | 有(在葉綠體中) | |
| 基於視紫紅質的能量代謝 | 有 | 有 | 無 | |
| 化能無機營養(Fe, S, H2) | 有 | 有 | 無 | |
| 氣泡 | 有 | 有 | 無 | |
| 聚-β-羥基脂肪酸作為碳儲存顆粒 | 有 | 有 | 無 | |
| 在80℃以上生長 | 有 | 有 | 無 | |
基本信息
- 中文名
- 三域系統
- 提出者
- 卡爾·烏斯(Carl Woese)
- 提出時間
- 1977年
- 類型
- 細胞生命形式分類