古菌
古菌
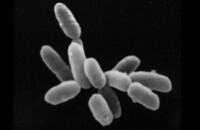
古菌可能是最古老的生命體,古菌一些奇特的生活習性和與此相關的潛在生物技術開發前景,長期以來一直吸引著許多人的注意。古菌常被發現生活於各種極端自然環境下,如大洋底部的高壓熱溢口、熱泉、鹽鹼湖等。
古菌(又稱古細菌、古生菌或太古生物、古核生物),是單細胞微生物,構成生物分類的一個域,或一個界。這些微生物屬於原核生物,它們與細菌有很多相似之處,即它們沒有細胞核與任何其他膜結合細胞器,同時另一些特徵相似於真核生物,比如存在重複序列與核小體。
古菌
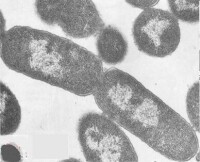
古菌的細胞結構與細菌不同,如古菌的細胞外膜就與細菌不同。
大多數古菌的細胞壁不含二氨基庚二酸(D-氨基酸)和胞壁酸,不受溶菌酶和內醯胺抗生素如青霉素的作用。革蘭氏陽性古菌的細胞壁含有各種複雜的多聚體,如產甲烷菌的細胞壁含假肽聚糖,甲烷八疊球菌和鹽球菌不含假肽聚糖,而含複雜聚多糖。革蘭氏陰性古菌沒有外膜,含蛋白質或糖蛋白亞基的表層,其厚度在20~40nm。甲烷葉菌屬、鹽桿菌屬和極端嗜熱的硫化葉菌屬、熱變形菌屬和熱網菌屬的細胞壁有糖蛋白;甲烷球菌屬、甲烷微菌屬、產甲烷菌屬和極端嗜熱的脫硫球菌屬有蛋白質壁,蛋白質呈酸性。
古菌在代謝過程中,有許多特殊的輔酶。古菌有5個類群,所以,它們的代謝呈多樣性。古菌中有異養型、自養型和不完全光合作用3種類型。
古菌
古菌利用二分裂、分裂和出芽進行無性繁殖;古菌不會進行減數分裂,所以擁有同樣的基因的一個種的古菌可能擁有不同的形態。古菌的細胞分裂被它們的細胞周期所控制;在細胞的染色體複製並分離后,細胞開始一分為二。雖說目前古菌中只有硫化葉菌的複製周期被闡明,但是這樣的細胞周期大體上類似於在細菌和真核生物的周期。和真核生物一樣,古菌的染色體也可以在多個位點(複製起點)開始用DNA聚合酶複製。然而古菌用於控制細胞分裂的蛋白(例如FtsZ,製造逐漸縮小的“Z環”以幫助細胞質分裂)和分離兩個子細胞的隔膜部分,這些與細菌的二分裂相似。
但與細菌、真核生物不同,目前未發現有古菌進行孢子生殖。一些嗜鹵鹽菌的種類可以進行表型轉換並生長成為不同的形態。這些形態包括擁有可以防止滲透壓休克出現的厚細胞壁,這使嗜鹵鹽菌可以在鹽度低的水中存活。這些古菌特徵不是生殖結構,但是它們可以幫助古菌在新的環境下生存。
大多數古菌生活在極端環境,如鹽分高的湖泊水中,極熱、極酸和絕對厭氧的環境,有的在極冷的環境生存。
按照古菌的生活習性和生理特性,古菌可分為三大類型:產甲烷菌、嗜熱嗜酸菌、極端嗜鹽菌。
從2001年起著手出版的《伯傑氏系統細菌學手冊》第二版中,將原核生物分為古菌域和細菌域。
根據16SrRNA序列分析,古菌域可分類為泉古菌門(界),廣古菌門(界),初古菌門(界),和納古菌門(界)。
隨著微生物分子生態學技術的發展和應用顯示,在中溫環境中存在著廣泛的大量未知的與泉古菌有密切聯繫的一類古菌,被稱之為中溫泉古菌(non-thermophilic Crenarchaeota)。隨著技術的發展發現諸多證據均不支持中溫泉古菌由嗜熱泉古菌進化而來的假設。因而專家建議將這些古菌(中溫泉古菌)劃分為一個新的門,成為古菌域的第三個主要類群一Thaumarchaeota(意譯為奇古菌門)。
大多數泉古菌門極端嗜熱、嗜酸,代謝硫。硫在厭氧呼吸中作為電子受體和無機營養的電子源。它們多生長在含硫地熱水或土壤中(如美國的黃石國家公園的富硫溫泉)。
熱變形菌綱(Thermoprotei)
暖球形菌目(Caldisphaerales)
暖球形菌科(Caldisphaeraceae)
暖球形菌屬(Caldisphaera)
除硫球菌目(Desulfurococcales)
除硫球菌科(Desulfurococcaceae)
酸葉菌屬(Acidilobus)
氣火菌屬(Aeropyrum)
除硫球菌屬(Desulfurococcus)
燃球菌屬(Ignicoccus)
燃球形菌屬(Ignisphaera)
葡萄熱菌屬(Staphylothermus)
施鐵特菌屬(Stetteria)
厭硫球菌屬(Sulfophobococcus)
熱盤菌屬(Thermodiscus)
熱球形菌屬(Thermosphaera)
熱網菌科(Pyrodictiaceae)
超熱菌屬(Hyperthermus)
熱網菌屬(Pyrodictium)
火葉菌屬(Pyrolobus)
硫化葉菌目(Sulfolobales)
硫化葉菌科(Sulfolobaceae)
喜酸菌屬(Acidianus)
除硫葉菌屬(Desulfurolobus)
生金球形菌屬(Metallosphaera)
憎葉菌屬(Stygiolobus)
硫化葉菌屬(Sulfolobus)
硫磺球形菌屬(Sulfurisphaera)
硫磺球菌屬(Sulfurococcus)
熱變形菌目(Thermoproteales)
熱絲菌科(Thermofilaceae)
熱絲菌屬(Thermofilum)
熱變形菌科(Thermoproteaceae)
暖枝菌屬(Caldivirga)
火棒菌屬(Pyrobaculum)
熱分支菌屬(Thermocladium)
熱變形菌屬(Thermoproteus)
火山鬃菌屬(Vulcanisaeta)
古丸菌綱(Archaeoglobi)
古丸菌目(Archaeoglobales)
古丸菌科(Archaeoglobaceae)
古丸菌屬(Archaeoglobus)
鐵丸菌屬(Ferroglobus)
地丸菌屬(Geoglobus)
鹽桿菌綱(Halobacteria)
鹽桿菌目(Halobacteriales)
鹽桿菌科(Halobacteriaceae)
適鹽菌屬(Haladaptatus)
鹽鹼球菌屬(Halalkalicoccus)
鹽盒菌屬(Haloarcula)
鹽桿菌屬(Halobacterium)
鹽棒菌屬(Halobaculum)
鹽二型菌屬(Halobiforma)
鹽球菌屬(Halococcus)
富鹽菌屬(Haloferax)
鹽幾何菌屬(Halogeometricum)
鹽微菌屬(Halomicrobium)
鹽惰菌屬(Halopiger)
鹽盤菌屬(Haloplanus)
鹽方菌屬(Haloquadratum)
鹽棍菌屬(Halorhabdus)
鹽紅菌屬(Halorubrum)
鹽簡菌屬(Halosimplex)
鹽池棲菌屬(Halostagnicola)
鹽陸生菌屬(Haloterrigena)
鹽長命菌屬(Halovivax)
鈉白菌屬(Natrialba)
鈉線菌屬(Natrinema)
鹽鹼桿菌屬(Natronobacterium)
鹽鹼球菌屬(Natronococcus)
鹽鹼湖菌屬(Natronolimnobius)
鹽鹼單胞菌屬(Natronomonas)
鹽鹼紅菌屬(Natronorubrum)
甲烷桿菌綱(Methanobacteria)
甲烷桿菌目(Methanobacteriales)
甲烷桿菌科(Methanobacteriaceae)
甲烷桿菌屬(Methanobacterium)
甲烷短桿菌屬(Methanobrevibacter)
甲烷球形菌屬(Methanosphaera)
甲烷熱桿菌屬(Methanothermobacter)
甲烷熱菌科(Methanothermaceae)
甲烷熱菌屬(Methanothermus)
甲烷球菌綱(Methanococci)
甲烷球菌目(Methanococcales)
甲烷暖球菌科(Methanocaldococcaceae)
甲烷暖球菌屬(Methanocaldococcus)
甲烷炎菌屬(Methanotorris)
甲烷球菌科(Methanococcaceae)
甲烷球菌屬(Methanococcus)
甲烷熱球菌屬(Methanothermococcus)
甲烷微菌綱(Methanomicrobia)
甲烷微菌目(Methanomirobiales)
甲烷粒菌科(Methanocorpusculaceae)
甲烷粒菌屬(Methanocorpusculum)
甲烷微菌科(Methanomicrobiaceae)
甲烷囊菌屬(Methanoculleus)
甲烷泡菌屬(Methanofollis)
產甲烷菌屬(Methanogenium)
甲烷裂葉菌屬(Methanolacinia)
甲烷微菌屬(Methanomicrobium)
甲烷盤菌屬(Methanoplanus)
甲烷螺菌科(Methanospirillaceae)
甲烷螺菌屬(Methanospirillum)
甲烷火菌綱(Methanopyri)
甲烷火菌目(Methanopyrales)
甲烷火菌科(Methanopyraceae)
甲烷火菌屬(Methanopyrus)
熱球菌綱(Thermococci)
熱球菌目(Thermococcales)
熱球菌科(Thermococcaceae)
古老球菌屬(Palaeococcus)
火球菌屬(Pyrococcus)
熱球菌屬(Thermococcus)
熱原體綱(Thermoplasmata)
熱原體目(Thermoplasmatales)
鐵原體科(Ferroplasmaceae)(註:正確形式應為Ferroplasmataceae)
鐵原體屬(Ferroplasma)
嗜苦菌科(Picrophilaceae)
嗜苦菌屬(Picrophilus)
熱原體科(Thermoplasmataceae)
熱原體屬(Thermoplasma)
分類疑物種
甲烷礫菌屬(Methanocalculus)
甲烷繩菌屬(Methanolinea)
甲烷八疊球菌目(Methanosarcinales)
甲烷鬃菌科(Methanosaetaceae)
甲烷鬃菌屬(Methanosaeta)
甲烷發菌屬(Methanothrix)
甲烷八疊球菌科(Methanosarcinaceae)
鹽甲烷球菌(Halomethanococcus)
甲烷微球菌屬(Methanimicrococcus)
甲烷類球菌屬(Methanococcoides)
甲烷鹽菌屬(Methanohalobium)
甲烷嗜鹽菌屬(Methanohalophilus)
甲烷葉菌屬(Methanolobus)
甲烷食甲基菌屬(Methanomethylovorans)
甲烷咸菌屬(Methanosalsum)
甲烷八疊球菌屬(Methanosarcina)
甲熱球菌科(Methermicoccaceae)
甲熱球菌屬(Methermicoccus)
科未定
熱裸單胞菌屬(Thermogymnomonas)
該類菌種目前只能用熒光原位雜交技術檢測證實其存在,實驗室中尚有未被培養的微生物(VBNC,viable but nonculturable microorganisms)。
初古菌屬(Korarchaeum)
2005年德國科學家在北冰洋底發現了一種寄生古菌中的最古老、最簡單、最微小的細菌騎火球納米古生菌(Nanoarchaeum equitans),只有大腸桿菌的160分之一(與天花病毒大小相當)、擁有500萬個鹼基(比最小支原體還少8萬個)。
納古菌屬(Nanoarchaeum)
該類菌種是最新發現的極端物種。
餐古菌目(Cenarchaeales)
餐古菌科(Cenarchaeaceae)
餐古菌屬(Cenarchaeum)
亞硝化暖菌目(Nitrosocaldales)
亞硝化暖菌科(Nitrosocaldaceae)
亞硝化暖菌屬(Nitrosocaldus)
亞硝化侏儒菌目(Nitrosopumilales)
亞硝化侏儒菌科(Nitrosopumilaceae)
亞硝化侏儒菌屬(Nitrosopumilus)
“古細菌”這個概念是1977年由卡爾·沃斯和George Fox提出的,原因是它們在16SrRNA的系統發生樹上和其它原核生物的區別。這兩組原核生物起初被定為古細菌(Archaebacteria)和真細菌(Eubacteria)兩個界或亞界。Woese認為它們是兩支根本不同的生物,於是重新命名其為古菌(Archaea)和細菌(Bacteria),這兩支和真核生物(Eukarya)一起構成了生物的三域系統。
如果將地球約46億年的年齡比作一年,那麼古菌早在3月20日就出現了,而人類誕生不過是12月31日的事。它們多生長於極端環境,如熱泉、高壓的海底火山口、鹽湖等。
人們對古菌的興趣並非始於1970年代。事實上,在我們這個星球上,古菌代表著生命的極限,確定了生物圈的範圍。例如,一種叫做熱網菌(Pyrodictium)的古菌能夠在高達113℃的溫度下生長。這是迄今為止發現的最高生物生長溫度。近年來,利用分子生物學方法,人們發現,古菌還廣泛分佈於各種自然環境中,土壤、海水、沼澤地中均生活著古菌。
目前,可在實驗室培養的古菌主要包括三大類:產甲烷菌、極端嗜熱菌和極端嗜鹽菌。產甲烷菌生活於富含有機質且嚴格無氧的環境中,如沼澤地、水稻田、反芻動物的反芻胃等,參與地球上的碳素循環,負責甲烷的生物合成;極端嗜鹽菌生活於鹽湖、鹽田及鹽腌製品表面,它能夠在鹽飽和環境中生長,而當鹽濃度低於10%時則不能生長;極端嗜熱菌通常分佈於含硫或硫化物的陸相或水相地質熱點,如含硫的熱泉、泥潭、海底熱溢口等,絕大多數極端嗜熱菌嚴格厭氧,在獲得能量時完成硫的轉化。
儘管生活習性大相徑庭,古菌的各個類群卻有共同的、有別於其他生物的細胞學及生物化學特徵。例如,古菌細胞膜含由分枝碳氫鏈與D型磷酸甘油,以醚鍵相連接而成的脂類,而細菌及真核生物細胞膜則含由不分枝脂肪酸與L型磷酸甘油,以酯鍵相連接而成的脂類。細菌細胞壁的主要成分是肽聚糖,而古菌細胞壁不含肽聚糖。
有趣的是,雖然與細菌相似,古菌染色體DNA呈閉合環狀,基因也組織成操縱子(操縱子為原核生物基因表達和調控的基本結構單位,生物活性相關的基因常以操縱子的結構形式協調基因表達的開啟和關閉),但在DNA複製、轉錄、翻譯等方面,古菌卻具有明顯的真核特徵:採用非甲醯化甲硫氨醯tRNA作為起始tRNA,啟動子、轉錄因子、DNA聚合酶、RNA聚合酶等均與真核生物的相似。
比較生物化學的研究結果表明,古菌與細菌有著本質的區別,這種區別與兩者表現在系統發育學方面親緣關係的疏遠是一致的。
地球上究竟有幾種生命形式?當亞里士多德建立生物學時,他用二分法則將生物分為動物和植物。顯微鏡的誕生使人們發現了肉眼看不見的細菌。細菌在細胞結構上與動植物的最根本差別是,動植物細胞內有細胞核,遺傳物質DNA主要儲存於此,而細菌則沒有細胞核,DNA遊離於細胞質中。由於動物與植物的差別小於它們與細菌的差別,沙東(E.Chatton)於1937年提出了生物界新的二分法則,即生物分為含細胞核的真核生物和不含細胞核的原核生物。動植物屬於真核生物,而細菌屬於原核生物。
1859年達爾文發表《物種起源》以後,生物學家便開始建立基於進化關係而非表型相似性的分類系統,即所謂系統發育分類系統。可是,由於缺乏化石記錄,這種分類方法長期未能有效運用於原核生物的分類。1970年代,隨著分子生物學的發展,伍斯終於在這方面獲得了意義重大的突破。
在漫長的進化過程中,每種生物細胞中的信息分子(核酸和蛋白質)的序列均不斷發生著突變。許多信息分子序列變化的產生在時間上是隨機的,進化速率相對恆定,即具有時鐘特性。因此,物種間的親緣關係可以用它們共有的某個具有時鐘特性的基因或其產物(如蛋白質)在序列上的差別來定量描述。這些基因或其產物便成了記錄生物進化歷程的分子記時器(chronometer)。顯然,這種記錄生物系統發育歷程的分子記時器應該廣泛分佈於所有生物之中。基於這一考慮,伍斯選擇了一種名為小亞基核糖體核酸(SSU rRNA)的分子,作為分子記時器。這種分子是細胞內蛋白質合成機器——核糖體的一個組成部分,而蛋白質合成又是幾乎所有生物生命活動的一個重要方面。因此,把SSU rRNA分子作為分子記時器是合適的。
在比較了來自不同原核及真核生物的SSU rRNA序列的相似性后,伍斯發現原先被認為是細菌的甲烷球菌代表著一種既不同於真核生物,也不同於細菌的生命形式。考慮到甲烷球菌的生活環境可能與生命誕生時地球上的自然環境相似,伍斯將這類生物稱為古細菌。據此,伍斯於1977年提出,生物可分為三大類群,即真核生物、真細菌和古細菌。基於SSU rRNA分析結果的泛系統發育(進化)樹隨後誕生了。
進一步的研究表明,進化樹上的第一次分叉產生了真細菌的一支和古細菌/真核生物的一支,古細菌和真核生物的分叉發生在後。換句話說,古細菌比真細菌更接近真核生物。
據此,1990年伍斯提出了三域分類學說:生物分為真核生物、真細菌和古細菌三域,域被定義為高於界的分類單位。為突出古細菌與真細菌的區別,伍斯將古細菌更名為古菌。真細菌改稱細菌。三域學說使古菌獲得了與真核生物和細菌等同的分類學地位。
自誕生之日起,伍斯的三域學說便遭到部分人,特別是微生物學領域外的人反對。反對者堅持認為:原核與真核的區分是生物界最根本的、具有進化意義的分類法則;與具有豐富多樣性表型的真核生物相比,古菌與細菌的差異遠沒有大到需要改變二分法則的程度。但在詹氏甲烷球菌基因組序列測定完成前的近20年中,採用多種分子記時器進行的系統發育學研究一再證明,古菌是一種獨特的生命形式。
儘管對古菌已有了上述認識,當人們第一次面對詹氏甲烷球菌的全基因組序列時,還是大吃了一驚。詹氏甲烷球菌共有1738個基因,其中人們從未見過的基因竟佔了56%!相比之下,在這之前完成測序的流感嗜血菌(Haemophilus influenzae)和生殖道枝原體(Mycoplasma genitalium)基因組中未知基因僅佔20%左右。於是人們終於在基因組水平上認識到,古菌是一種嶄新的生命形式。
更有趣的是,詹氏甲烷球菌基因組中佔總數44%的那些功能或多或少已經知道的基因似乎勾勒出了古菌與另兩類生物之間的進化關係:古菌在產能、細胞分裂、代謝等方面與細菌相近,而在轉錄、翻譯和複製方面則與真核生物類似。換言之,一個生活在大洋底部熱溢口處的、習性古怪的微生物,在遺傳信息傳遞方面竟有著與人(而不是與人的消化道中細菌)相似的基因!在讚歎生命奇妙的同時,許多人開始歡呼三域學說的最終確立。美國《科學》周刊在把詹氏甲烷球菌基因組序列測定工作列為1996年度重大科學突破之一時宣稱,這一成果使圍繞三域學說的爭論差不多可以結束了。
就在古菌的懸念似乎行將消失時,接踵而來的新發現卻使人們重新陷入困惑之中。各類完整的微生物基因組序列一個接一個出現在人們輕點滑鼠便可查閱的資料庫中,在已發表的18種生物基因組序列中,古菌的佔了4個。採用更靈敏的方法對這些基因組(包括詹氏甲烷球菌基因組)進行分析,得到了令人吃驚的結果:詹氏甲烷球菌基因組中只有30%(不是先前估計的半數以上)的基因編碼目前未知的功能,這與細菌基因組相近。古菌的神秘性和獨特性因此減少了許多。
對三域學說更為不利的是,在詹氏甲烷球菌的那些可以推測功能的基因產物(蛋白質)中,44%具有細菌蛋白特徵,只有13%的像真核生物的蛋白質。在另一個古菌,嗜熱鹼甲烷桿菌(Methanobacterium thermoaotutrophicum)的基因組中也有類似情況。因此,從基因組比較的數字上看,古菌與細菌間的差異遠小於古菌與真核生物間的差異,不足以說服三域學說的反對者。
更令人難以理解的是,利用同一生物中不同基因對該物種進行系統發育學定位常常會得到不同的結果。最近,一種能在接近沸點溫度下生長的細菌(Aquifex aeolicus)的基因組序列測定完成。對該菌的幾個基因進行的系統發育學研究表明:如果用參與細胞分裂調控的蛋白質FtsY作為分子記時器,該菌與伍斯進化樹上位於細菌分枝的一個土壤細菌——枯草芽孢桿菌相近;如果以一種參與色氨酸合成的酶為準,該菌應屬於古菌;而當比較該菌和其他生物的合成胞苷三磷酸(DNA的基本結構單位之一)的酶時,竟發現古菌不再形成獨立的一群。看來不同的基因似乎在訴說不同的進化故事。那麼,古菌還能是獨特的、統一的生命形式嗎?
屬於真核生物的啤酒酵母基因組序列測定完成後,三域學說遇到了更大危機。酵母細胞核基因中,與細菌基因有親緣關係的比與古菌有親緣關係的多一倍。有人還對在三種生命形式中都存在的34個蛋白質家族進行了分析,發現其中17個家族來源於細菌,只有8個顯示出古菌與真核生物的親緣關係。
如果伍斯進化樹正確、古菌與真核生物在進化歷程中的分歧晚於兩者與細菌的分歧的話,那麼怎樣才能解釋上面這些結果呢?
根據細胞進化研究中流行的內共生假說,真核細胞細胞器(線粒體、葉綠體)的產生源於細菌與原真核生物在進化早期建立的內共生關係。在這種關係中,真核細胞提供穩定的微環境,內共生體(細菌)則提供能量,久而久之,內共生體演變為細胞器。真核生物細胞核中一部分源於細菌的基因可能來自線粒體,這些為數不多的基因通常編碼重新運回線粒體的蛋白質分子。可是,現在發現許多源於細菌的核基因編碼那些在細胞質、而不是線粒體中起作用的蛋白質。那麼,這些基因從何而來呢?顯然,內共生假說已不足以挽救伍斯進化樹。
不過,伍斯進化樹也不會輕易倒下,支撐它的假說依然很多。最近,有人提出了新版的“基因水平轉移”假說。根據這個假說,基因組的雜合組成是進化過程中不同譜系間發生基因轉移造成的。一種生物可以採用包括吞食等方式獲得另一種、親緣關係也許很遠的生物的基因。伍斯推測,始祖生物在演化形成細菌、古菌和真核生物三大譜系前,生活於可以相互交換基因的“公社”中,來自這個“史前公社”的生物可能獲得了不同的基因遺產。這一切使得進化樹難以枝杈分明。不過,伍斯相信,基於SSUrRNA的進化樹在總體上是正確的,三種生命形式是存在的。
三年前詹氏甲烷球菌基因組序列的發表,似乎預示著一場延續了20多年的、關於地球上到底有幾種生命形式的爭論的終結。古菌似乎被認定為生命的第三種形式。如今,僅僅過了三年,即使最樂觀的人都無法預料伍斯進化樹的命運。這場仍在繼續的爭論中,儘管古菌的分類地位遭到質疑,但古菌這一生命形式的獨特性依然得到不同程度的肯定。
目前,古菌研究正在世界範圍內升溫,這不僅因為古菌中蘊藏著遠多於另兩類生物的、未知的生物學過程和功能,以及有助於闡明生物進化規律的線索,而且因為古菌有著不可估量的生物技術開發前景。古菌已經一次又一次讓人們吃驚,可以肯定,在未來的歲月中,這群獨特的生物將繼續向人們展示生命的無窮奧秘。

基本信息
- 中文名
- 古菌
- 別名
- 古細菌、太古菌或太古生物
- 界
- 古細菌界
- 門
- 泉古菌門,廣古菌門
- 分布區域
- 大洋底部的高壓熱溢口、熱泉、鹽鹼湖等
- 繁殖方式
- 二分裂、芽殖
- 形態
- 球形、桿狀、螺旋形、耳垂形等
- 細胞結構
- 細胞壁不含二氨基庚二酸
- 生活習性
- 生活在極端環境
- 分類
- 產甲烷菌、嗜熱嗜酸菌、極端嗜鹽菌
- 域
- 古菌域