轉錄因子
轉錄因子
徠轉錄因子(Transcription factors )。
真核生物轉錄起始過程十分複雜,往往需要多種蛋白因子的協助,轉錄因子與RNA聚合酶Ⅱ形成轉錄起始複合體,共同參與轉錄起始的過程。根據轉錄因子的作用特點可分為二類;第一類為普遍轉錄因子,它們與RNA聚合酶Ⅱ共同組成轉錄起始複合體時,轉錄才能在正確的位置開始。除TFⅡD以外,還發現TFⅡA,TFⅡB,TFⅡF,TFⅡE,TFⅡH等,它們在轉錄起始複合體組裝的不同階段起作用。第二類轉錄因子為組織細胞特異性轉錄因子,這些TF是在特異的組織細胞或是受到一些類固醇激素\生長因子或其它刺激后,開始表達某些特異蛋白質分子時,才需要的一類轉錄因子。

RNA轉錄過程
人類金屬硫基因調節區
轉錄因子的結合位點(transcription factor binding site,TFBS)是轉錄因子調節基因表達時,與基因模板鏈結合的區域。按照常識,轉錄因子(transcription factor)的結合位點一般應該分佈在基因的前端,但是,新的研究發現,人21和22號染色體上,只有22%的轉錄因子結合位點分佈在蛋白編碼基因的5'端。
真核生物在轉錄時往往需要多種蛋白質因子的協助。一種蛋白質是不是轉錄結構的一部分往往是通過體外系統看它是否是轉錄起始所的。一般這些促成轉錄起始所需的轉錄結構包括三個重要的組成部分:
某些轉錄因子能與RNA聚合酶結合形成起始複合物,但不組成遊離聚合酶的成分。這些因子可能是所有啟動子起始轉錄所必需的,但亦可能僅是譬如說轉錄終止所必需的。但是,在這一類因子中,要嚴格區分開哪些是RNA聚合酶的亞基,哪些僅是輔助因子,是很困難的。
某些轉錄因子僅與其靶啟動子中的特異順序結合。如果這些順序存在於啟動子中,則這些順序因子是一般轉錄結構的一部分。如果這些順序僅存在於某些種類的啟動子中,則識別這些順序的因子也只是在這些特異啟動子上起始轉錄必需的。
黑腹果蠅的RNA聚合酶需要至少兩個轉錄因子方能在體外起始轉錄。其中一個是B因子,它與含TATA盒的部位結合。人的因子TFⅡD亦和類似的部位結合。同樣,CTF(CAAT結合因子)則與腺病毒的主要晚期啟動子中與CAAT盒同源的部位相結合。結合在上游區的另一個轉錄因子是USF(亦稱MLTF),則可以識別腺病毒晚期啟動子中靠近-55的順序。轉錄因子Sp1則能和GC盒相結合。在SC40啟動子中有多個GC盒,位於-70到-110之間。它們均能和Sp1相結合。然而含有GC盒的不同的DNA順序與Sp1的親和力卻各不相同。可見GC盒兩側的順序對Sp1-GC盒的結合究竟如何能影響轉錄。有時候需要幾個轉錄因子才能起始轉錄。例如胞苷激酶的啟動子需要Sp1與GC盒結合和CTF與CAAT盒結合;腺病毒晚期啟動子需要TFⅡD與TATA盒結合和USF與其鄰近部位相結合。以上所述的因子是一般轉錄都需要的,似乎並沒有什麼調節功能。另一些轉錄因子則可以調控一組特殊基因的轉錄。熱休克基因就是一個很好的例子。真核生物的熱休克基因在轉錄起始點的上游15bp處有一個共同順序。HSTF因子僅在熱休克細胞中有活性。它與包括熱休克共同順序在內的一段DNA相結合,所以這個因子的激活可以引起約包括20個基因的一組基因起始轉錄。在這裡,轉錄因子和RNA聚合酶Ⅱ之間關係很類似細菌的σ因子與核心酶之間的關係。
轉錄因子是一種具有特殊結構、行使調控基因表達功能的蛋白質分子,也稱為反式作用因子。植物中的轉錄因子分為二種,一種是非特異性轉錄因子,它們非選擇性地調控基因的轉錄表達,如大麥 (Hordeum vulgare) 中的HvCBF2 (C-repeat/DRE binding factor 2) (Xue et al., 2003)。還有一種稱為特異型轉錄因子,它們能夠選擇性調控某種或某些基因的轉錄表達。典型的轉錄因子含有DNA結合區 (DNA-binding domain)、轉錄調控區 (activation domain)、寡聚化位點(oligomerization site) 以及核定位信號 (nuclear localization signal) 等功能區域。這些功能區域決定轉錄因子的功能和特性 (Liu et al., 1999)。DNA結合區帶共性的結構主要有:1)HTH 和 HLH 結構:由兩段α-螺旋夾一段β-摺疊構成,α-螺旋與β-摺疊之間通過β-轉角或成環連接,即螺旋-轉角-螺旋結構和螺旋-環-螺旋結構。2)鋅指結構:多見於 TFIII A 和類固醇激素受體中,由一段富含半胱氨酸的多肽鏈構成。每四個半光氨酸殘基或組氨酸殘基螯合一分子 Zn2+ ,其餘約 12-13 個殘基則呈指樣突出,剛好能嵌入 DNA 雙螺旋的大溝中而與之相結合。3)亮氨酸拉鏈結構:多見於真核生物 DNA 結合蛋白的 C 端,與癌基因表達調控有關。由兩段α - 螺旋平行排列構成,其α - 螺旋中存在每隔 7 個殘基規律性排列的亮氨酸殘基,亮氨酸側鏈交替排列而呈拉鏈狀,兩條肽鏈呈鉗狀與 DNA 相結合。
同一家族的轉錄因子之間的區別主要在轉錄調控區。
轉錄調控區包括轉錄激活區 (transcription activation domain) 和轉錄抑制區 (transcription repression domain) 二種。近年來,轉錄的激活區被深入研究。它們一般包含DNA結合區之外的30-100個氨基酸殘基,有時一個轉錄因子包含不止一個轉錄激活區。如控制植物儲藏蛋白基因表達的VP1和PvALF轉錄因子,它們的N-末端酸性氨基酸保守序列都具有轉錄激活能力,與酵母轉錄因子GCN4和病毒轉錄因子的VP16的酸性氨基酸轉錄激活區有較高同源性 (Bobb et al., 1996)。典型的植物轉錄因子激活區一般富含酸性氨基酸、脯氨酸或谷氨醯胺等,如GBF (G-box binding factor) 含有的GCB盒 (GBF conserved box) 激活結構域 (lunwen114 and Bevan, 1998)。
也是轉錄因子調控表達的重要位點,但是對其作用機理研究尚不深入。可能的作用方式有三種:1)與啟動子的調控位點結合,阻止其它轉錄因子的結合;2)作用於其它轉錄因子,抑制其它因子的作用;3)通過改變DNA的高級結構阻止轉錄的發生。
轉錄因子必須在核內作用,才能起到調控表達的目的。因此,轉錄因子上的核定位序列是其重要的組成部分。一般一個或多個核定位序列在轉錄因子中不規則分佈,同時也存在不含核定位序列的轉錄因子,它們通過結合到其它轉錄因子上進入細胞核。核定位序列一般是轉錄因子中富含精氨酸和賴氨酸殘基的區段。目前,水稻中的GT-2、西紅柿中的HSFA1-2、玉米的O2和碗豆的PS-IAA4和6等轉錄因子中的核定位序列都已被鑒定 (Boulikas, 1994; Dehesh et al., 1995; Lyck et al., 1997; Varagona et al., 1992; Abel and Theologis, 1995)。
絕大多數轉錄因子結合 DNA前需通過蛋白質-蛋白質相互作用形成二聚體或多聚體。所謂二聚體化就是指兩分子單體通過一定的結構域結合成二聚體,它是轉錄因子結合DNA時最常見的形式。由同種分子形成的二聚體稱同二聚體,異種分子間形成的二聚體稱異二聚體。這種多聚體的形成是轉錄因子上的寡聚化位點 (oligomerization site) 相互作用的結果,寡聚化位點的氨基酸序列很保守,大多與DNA結合區相連並形成一定的空間構象。除二聚化或多聚化反應,還有一些調節蛋白不能直接結合DNA,而是通過蛋白質-蛋白質相互作用間接結合DNA,調節基因轉錄,這樣就形成了一個表達調控的複合物。
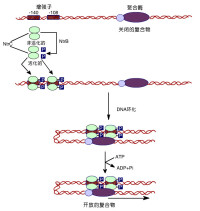
真核細胞轉錄因子
目前,人工轉錄因子(Artificial Transcription Factor,ATF)的構建已用於轉錄因子的生物學功能研究中起到重要作用。ATF是指將不同的DNA結合結構域與效應結構域組合在一起,人為地構建具有新的序列特異性與作用效果的轉錄因子,在基礎研究、藥物設計以及基因治療等領域得到了廣泛研究,目前研究較多的ATF主要為C2H2型鋅指結構。

基本信息
- 中文名
- 轉錄因子
- 外文名
- Transcription factors
- 生物名詞
- 一群能與基因5`端上游特定序列
- 實質
- 蛋白質
- 類別
- 生物名詞