細胞核
細胞遺傳與代謝的調控中心
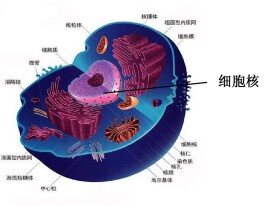
細胞核(nucleus)是真核細胞內最大、最重要的細胞結構,是細胞遺傳與代謝的調控中心,是真核細胞區別於原核細胞最顯著的標誌之一(極少數真核細胞無細胞核,如哺乳動物的成熟的紅細胞,高等植物成熟的篩管細胞等)。(初中老教材、高中教材或一些國外教材認為細胞核不是細胞器,大學細胞生物學則認為是細胞器,這裡以大學教材為準),它主要由核膜(nuclear membrane)、染色質(chromatin)、核仁(nucleolus)、核基質(nuclear matrix)等組成。
細胞核是細胞內遺傳信息的儲存、複製和轉錄的主要場所。它是英國科學家布朗於1831年發現並命名的。大多呈球形或橢圓形。通常一個,也有兩個或多個的。借雙層多孔的核臘與細胞質分隔。核內含有核液、染色質(或染色體)和核仁。
細胞核是存在於真核細胞中的封閉式膜狀胞器,內部含有細胞中大多數的遺傳物質,也就是DNA。這些DNA與多種蛋白質,如組織蛋白複合形成染色質。而染色質在細胞分裂時,會濃縮形成染色體,其中所含的所有基因合稱為核基因。細胞核的作用,是維持基因的完整性,並藉由調節基因表現來影響細胞活動。
細胞核的主要構造為核膜,是一種將細胞核完全包覆的雙層膜,可使膜內物質與細胞質、以及具有細胞骨架功能的網狀結構核纖層分隔開來。由於多數分子無法直接穿透核膜,因此需要核孔作為物質的進出通道。這些孔洞可讓小分子與離子自由通透;而如蛋白質般較大的分子,則需要載體蛋白的幫助才能通過。核運輸是細胞中最重要的功能;基因表現與染色體的保存,皆有賴於核孔上所進行的輸送作用。
細胞核內不含有任何其他膜狀的結構,但也並非完全均勻,其中存在許多由特殊蛋白質、RNA以及DNA所複合而成的次核體。而其中受理解最透徹的是核仁,此結構主要參與核內RNA 的合成。RNA是核糖體的主要成分。核糖體在核仁中產出之後,會進入細胞質進行mRNA的轉譯。
細胞核是最早發現的細胞器,由弗朗茲·鮑爾(Franz Andreas Bauer)在1802年對其進行最早的描述。到了1831年,蘇格蘭植物學家羅伯特·布朗又在倫敦林奈學會的演講中,對細胞核做了更為詳細的敘述。布朗以顯微鏡觀察蘭花時,發現花朵外層細胞有一些不透光的區域,並稱其為“areola”或“nucleus”。不過他並未提出這些構造可能的功用。馬蒂亞斯·許萊登在1838年提出一項觀點,認為細胞核能夠生成細胞,並稱這些細胞核為“細胞形成核”(Cytoblast)。他也表示自己發現了組成於“細胞形成核”周圍的新細胞。不過弗朗茲·邁恩對此觀念強烈反對,他認為細胞是經由分裂而增殖,並認為許多細胞並沒有細胞核。由細胞形成核作用重新生成細胞的觀念,與羅伯特·雷馬克及魯道夫·菲爾紹的觀點衝突,他們認為細胞是單獨由細胞所生成。至此,細胞核的機能仍未明了。
在1876到1878年間,奧斯卡·赫特維希的數份有關海膽卵細胞受精作用的研究顯示,精子的細胞核會進到卵子的內部,並與卵子細胞核融合。首度闡釋了生物個體由單一有核細胞發育而成的可能性。這與恩斯特·海克爾的理論不同,海克爾認為物種會在胚胎髮育時期重演其種系發生歷程,其中包括從原始且缺乏結構的黏液狀“無核裂卵”(Monerula),一直到有核細胞產生之間的過程。因此精細胞核在受精作用中的必要性受到了漫長的爭論。赫特維希後來又在其他動物的細胞,包括兩棲類與軟體動物中確認了他的觀察結果。而愛德華·施特拉斯布格也從植物得到相同結論。這些結果顯示了細胞核在遺傳上的重要性。1873年,奧古斯特·魏斯曼提出了一項觀點,認為母系與父系生殖細胞在遺傳上具有相等的影響力。到了20世紀初,有絲分裂得到了觀察,而孟德爾定律也重新見世,這時候細胞核在攜帶遺傳訊息上的重要性已逐漸明朗。
細胞核是細胞的控制中心,在細胞的代謝、生長、分化中起著重要作用,是遺傳物質的主要存在部位。儘管細胞核的形狀有多種多樣,但是它的基本結構卻大致相同,主要由核被膜、染色質、核骨架、核仁及核體組成。
絕大多數真核生物細胞中;
(1)原核細胞中沒有真正的細胞核(稱為擬核);(2)有的真核細胞中也沒有細胞核,如哺乳動物的成熟的紅細胞,高等植物成熟的篩管細胞等極少數的細胞。
大多呈球形或卵圓形,但也隨物種和細胞類型不同而有很大變化。
核的大小依物種不同而有所變化,例如:高等動物細胞核直徑一般在5~10微米,高等植物細胞核直徑一般在5~20微米,低等植物細胞核直徑一般在1~4微米。
一般一個:大多數生物體細胞中都是一個;
人的骨胳肌中的細胞核可達數百個。
在HE染色切片上,細胞核以其強嗜鹼性而成為細胞內最醒目的結構。由於它含有DNA--遺傳信息,因此,借DNA複製與選擇性轉錄,細胞核成為細胞增殖、分化、代謝等活動中關鍵環節之一。人體絕大多數種類的細胞具有單個細胞核,少數無核、雙核或多核。核的形態在細胞周期各階段不同,間期核的形態在不同細胞亦相差甚遠,但其結構都包括核被膜,染色質,核仁與核基質四部。
核被膜使細胞核成為細胞中一個相對獨立的體系,使核內形成一相對穩定的環境。同時,核被膜又是選擇性滲透膜,起著控制核和細胞質之間的物質交換作用。
核被膜(nuclear envelope)包裹在核表面,由基本平行的內膜、外膜兩層膜構成。兩層膜的間隙寬10~15nm,稱為核周隙(perinuclear cisterna),也稱核周腔。核被膜上有核孔(nuclear pore)穿通,占膜面積的8%以上。外核膜表面有核糖體附著,並與粗面內質網相續;核周隙亦與內質網腔相通,因此,核被膜也參與蛋白質合成。內核膜也參與蛋白質合成。內核膜的核質面有厚20~80nm的核纖層(fibrous lamina)是一層由細絲交織形成的緻密網狀結構。成分為中間纖維蛋白,稱為核纖層蛋白(lamin)。核纖層與細胞質骨架、核骨架連成一個整體,一般認為核纖層為核被膜和染色質提供了結構支架。核纖層不僅對核膜有支持、穩定作用,也是染色質纖維西端的附著部位。
核孔是直徑80~120nm 的圓形孔,核孔複合體稍大些,直徑為120~150nm,內、外核膜在孔緣相連續,孔內有環(annulus)與中心顆粒組成核孔複合體。環有16個球形亞單位,孔內、外線各有8個。從位於核孔中心的中心顆粒(又稱孔栓)放射狀發出細絲與16個亞單位相連。核孔所在處無核纖層。一般認為,水離子和核苷等小分子物質可直接通透核被膜;而RNA與蛋白質等大分子則經核孔出入核,但其出入方式尚不明了。顯然,核功能活躍的細胞核孔數量多。成熟的精子幾乎無核孔,而卵母細胞的核孔極其豐富,成為研究該結構的主要材料。
核被膜三個區域各自概要
— 核外膜:面向胞質,附有核糖體顆粒,與內質網相連。
— 核內膜:面向核質,表面上無核糖顆粒,膜上有特異蛋白,為核纖層提供結合位點。
— 核孔(nuclear pores):在內外膜的融合處形成環狀開口,又稱核孔複合體,直徑為50~100nm,一般有幾千個,核孔構造複雜,含100種以上蛋白質,並與核纖層緊密結合成為核孔複合體。是選擇性雙向通道。功能是選擇性的大分子出入(主動運輸),酶、組蛋白、mRNA、tRNA;存在電位差,對離子的出入有一定的調節控制作用。
是遺傳物質DNA和組蛋白在細胞間期的形態表現。在HE染色的切片上,染色質有的部分著色淺淡,稱為常染色質(euchromatin),是核中進行RNA轉錄的部位;有的部分呈強嗜鹼性。稱異染色質:(heterochromatin),是功能靜止的部分,故根據核的染色狀態可推測其功能活躍程度。電鏡下,染色質由顆粒與細絲組成,在常染色所部分呈稀疏,在異染色質則極為濃密。現已證明,染色質的基本結構為串珠狀的染色質絲。染色質的結構單體為核小體,直徑約10nm,相鄰以1.5~2.5nm的細絲相連,核心由4組組蛋白( H2A,H2B,H3,H4 )構成,DNA纏繞在核心的外周,核小體之間為連接DNA,上有H1,1個核小體上共有200個鹼基對,構成染色質絲的一個單位。是由DNA雙股螺族鏈規則重複地盤繞,形成大量核小體(nucleosome)。核小體為直徑約10nm的扁圓球形,核心由5種蛋白(H1、 H2A、H2B、H3、H4)各二分子組成;DNA盤繞核心1.75周,含140個鹼基對。DNA鏈於相鄰核小體間走行的部分稱連接段,含10~70個鹼基對,並有組蛋白H1附著。這種直徑約10nm的染色質絲在其進行RNA轉錄的部位是舒展狀態,即表現為常染色質;而未執行動能的部位則螺旋化,形成直徑約30nm的染色質纖維,即異染色質。人體細胞核中含46條染色質絲,其DNA鏈總長約1m,只有以螺旋化狀態才能被容納於直徑4~5μm的核中。
是形成核糖體前身的部位。大多數細胞可具有1~4個核仁。在合成蛋白旺盛的細胞,核仁多而大。光鏡下,核仁呈圓形,並因含大量rRNA而顯強嗜鹼性。電鏡下,核 仁由細絲成分、顆粒成分與核仁相隨染色質三部分構成。細絲成分與顆粒成分是rRNA與相關蛋白質的不同表現形式,二者常混合組成粗約60~80nm核仁絲,後者蟠曲成網架。通常認為,顆粒成分是核糖體亞基的前身,由細絲成分逐漸轉變而成,可通過核孔進入細胞質;核仁相隨染色質是編碼rRNA的DNA鏈的局部。人的第13、14、15、21和22對染色體的一端有圓形的隨體(satellite),通過隨體柄與染色體其它部分相連。隨體柄即為合成rRNA的基因位點,又稱核仁組織者區(nucleons organizer region),當其解螺旋進入功能狀態時即成為核仁相隨染色質,並進一步發展為核仁。理論上人體細胞可有10個核仁,但在其形成過程中往往互相融合,因此細胞中核仁一般少於4個。
核仁經常出現在間期細胞核中,它是勻質的球體,其形狀、大小、數目依生物種類,細胞形成和生理狀態而異。核仁的主要功能是進行核糖體RNA的合成和核糖體的形成。
染色質和染色體在化學成分上並沒有什麼不同,而只是分別處於不同的功能階段的不同的構型。染色質是指間期細胞內由DNA、組蛋白和非組蛋白及少量RNA組成的線形複合結構,是間期細胞遺傳物質存在形式。固定染色后,在光鏡下能看到細胞核中經許多或粗或細的長絲交織成網的物質,從形態上可以分為常染色質(euchromatin)和異染色質(heterochromatin)。常染色質呈細絲狀,是DNA長鏈分子展開的部分,非常纖細,染色較淡。異染色質呈較大的深染團塊,常附在核膜內面,DNA長鏈分子緊縮盤繞的部分。染色體是指細胞在有絲分裂或減數分裂過程中,由染色質縮聚而成的棒狀結構。
核骨架是由纖維蛋白構成的網架結構,其蛋白成分按道理說細胞質骨架有的,核骨架也應該有。但在核骨架中只發現有角蛋白和肌蛋白質成分,在某些原生動物核骨架中還發現含有微管。同時在核骨架中還有少量RNA,它對於維持核骨架三維網路結構的完整性是必需的。在進化趨勢看,核骨架組分是由多樣化走向單一,特化。
從其結構,我們可以得出細胞核的功能:控制細胞的遺傳,生長和發育。德國藻類學哈姆林的傘藻嫁接試驗驗證了細胞核是遺傳物質攜帶者。
細胞核是細胞的控制中心,一般說真核細胞失去細胞核后,很快就會死亡,但紅細胞失去核后還能生活120天;植物篩管細胞,失去核后,能活好幾年。
1.遺傳物質儲存和複製的場所。從細胞核的結構可以看出,細胞核中最重要的結構是染色質,染色質的組成成分是蛋白質分子和DNA分子,而DNA分子又是主要遺傳物質。當遺傳物質向後代傳遞時,必須在核中進行複製。所以,細胞核是遺傳物儲存和複製的場所。
2.細胞遺傳性和細胞代謝活動的控制中心。遺傳物質能經複製後傳給子代,同時遺傳物質還必須將其控制的生物性狀特徵表現出來,這些遺傳物質絕大部分都存在於細胞核中。所以,細胞核又是細胞遺傳性和細胞代謝活動的控制中心。例如,英國的克隆綿羊“多莉”就是將一隻母羊卵細胞的細胞核除去,然後,在這個去核的卵細胞中,移植進另一個母羊乳腺細胞的細胞核,最後由這個卵細胞發育而成的。“多莉”的遺傳性狀與提供細胞核的母羊一樣。這一實例充分說明了細胞核在控制細胞的遺傳性和細胞代謝活動方面的重要作用。
因此,對細胞核功能的較為全面的闡述應該是:細胞核是遺傳信息庫,是細胞代謝和遺傳的控制中心。
細胞間信息交流的方式多種多樣,有間接傳遞信息的,如內分泌腺細胞將(激素)分泌出來后,由(血液)運送到全身各處作用與遠處的靶細胞;有直接傳遞信息的,如(精子)和(卵細胞)的識別和結合則是通過兩個細胞(細胞膜)直接接觸,完成信息交流。
一般認為細胞識別的分子基礎是受體。受體主要指細胞膜中的(糖蛋白)它對細胞外信號分子的結合有特異性。
1837年10月,施萊登把自己的實驗結果和想法告訴了柏林大學解剖生理學家施旺,並特別指出細胞核在植物細胞發生中所起的重要作用。施旺立刻回想起自己曾在脊索細胞中看見過的同樣“器官”,並意識到如果能夠成功地證明脊索細胞中的細胞核起著在植物細胞發生中所起的相同作用。那麼,這個發現將是極其有意義的。
施旺從植物細胞與動物細胞結構上的相似性出發,在細胞水平上完成了二者的統一工作。1839年他發表了《關於動植物結構和生長相似性的顯微研究》一文。全文內容有三部分:第一部分描述了他以動物為對象的研究情況和結論;第二部分提出了證據,把自己的實驗結果與施萊登的研究結果作對比,表明動物和植物的基本結構單位都是細胞;第三部分總結了全部研究結果,提出了細胞學說,詳細闡明了細胞的理論。施旺把施萊登證實了的植物的基本結構是細胞的觀點推廣到了動物界,並指出動植物發育的共同普遍規律。這在生物學史上具有劃時代的意義。施旺指出:“細胞是有機體,整個動物和植物體乃是細胞的集合體,它們依照一定的規律排列在動植物體內。”
細胞核起源依然是一個未解之謎。迄今為止的學說主要有:共營模型(syntrophic model)、自演化模型(autogenous model)、病毒性真核生物起源模型(viral eukaryogenesis model)、外膜假說(exomembrane hypothesis)、壓縮和結構化假說(packing and structurization hypothesis),等等。
共營模型認為,與現代產甲烷古菌類似的某些古老的古菌,侵入並生活在類似於現代粘細菌的細菌體內,形成了早期的細胞核。古菌與真核生物在特定蛋白質(如組蛋白)基因的相似性被認為是支持以古菌為基礎的細胞核起源理論的證據。
自演化模型認為原真核(proto-eukaryotic)細胞直接自細菌演化而來,並不需要通過內共生。證據來自一類專性好氧菌——浮黴菌(Planctomycete),它們具有清晰的胞內膜結構,其中,有一種稱之為Gemmata obscuriglobus的出芽菌,其染色質被雙層的核膜所包裹,類似於真核生物的核的結構,而斯氏小梨形菌(Pirellula staleyi)的核被單層的細胞質內膜ICM所包裹。但是,這一模型並未進一步解釋核實如何形成的。
病毒性真核生物起源模型認為,病毒感染原核生物導致了膜結合的細胞核與其他真核生物特徵的產生。證據是真核生物和病毒在大分子結構上存在一定相似性,譬如,線性DNA鏈、mRNA的加帽,以及與蛋白質的緊密結合(病毒的外套膜類似於組蛋白)。該假說的其中一種觀點認為,吞噬作用形成了早期的細胞“捕食者”,並隨之演化出細胞核。
外膜假說認為,細胞核是起源自演化出第二層外細胞膜的單個早期細胞,而包裹原來細胞的內膜則轉變成了核膜,並逐漸演化出精巧的核孔結構,以便於將內部(如核糖體亞基)合成的物質送出核外。
壓縮與結構化假說認為,細胞核源自原核細胞基因組的大型化(包括DNA的複製錯誤或多倍化、側向基因轉移方式、內共生融合等)。核的成型及有絲分裂的出現主要是為了滿足將巨大的DNA分子準確地分配到子代中去的需求,因此,如何將長鏈DNA有效地壓縮(藉助組蛋白)成若干染色體以及如何將多個染色體同時分離(藉助紡錘體)是核演化的關鍵。從原核生物到真核生物,基因組的DNA總量大約增加了3.5個數量級,這與現代真核生物的DNA壓縮比(packing ratio)驚人地一致。包括核膜在內的細胞內膜系統就是為了實現對複雜生化系統進行秩序化管控,或者說,秩序化是通過細胞內部的模塊化得以實現的。
基本信息
- 中文名
- 細胞核
- 外文名
- nucleus
- 組成部分
- 核膜,核骨架,核仁等
- 作用
- 是細胞遺傳與代謝的調控中心
- 發現時間
- 1802年