核膜
核膜
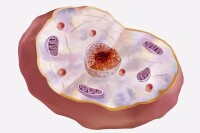
核膜是位於真核生物的核與細胞質交界處的雙層結構膜。核膜對核內外物質的交通有高度選擇性,控制細胞核內外物質交換運輸和信息傳輸。
核膜
核膜外層的外表面附有核糖體顆粒。有的細胞中,外膜與粗面內質網膜相連續,因為內質網膜與質膜是連續的,所以核膜間隙似乎與細胞外相通。
核膜上還有許多散在的孔,稱為核孔,在核孔周圍,核膜的內層與外層相連。核孔是核與細胞質進行物質交換的孔道。
核膜並不是完全連續的,有許多部位內外膜互相連接,形成穿過核膜的核孔。(註:無法用光學顯微鏡看見核膜)
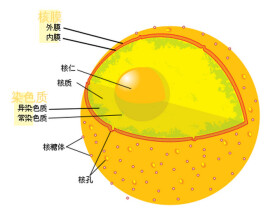
核膜有典型的脂雙層結構,以分佈為標準將其定為外膜和內膜區,膜間存在小的核孔。核膜的穩定性依靠兩層中間絲的網狀結構來維持:內部網路在內核膜上形成核層。外部形成較鬆散的網路以提供外部支持。
外膜與內質網的一部分相連接,但在核膜中的蛋白質濃度卻高於內質網腔中。核膜的這種基本結構,可因生物種類的不同而異。例如,綠藻類的角絲鼓藻只有一層核膜,但在變形蟲和某種脊椎動物的細胞中,在核膜內側則有第三層膜結構,即呈三層結構。另外,渦鞭蟲類中的夜光蟲有雙層膜結構,但核膜上無核孔。在高等真核生物的有絲分裂前期,核膜變成與小泡和內質網不能區分的幾個斷片。但到分裂末期,核膜在子染色體群表面重新形成,成為子核的核膜。另一方面,已知許多低等真核生物經過分裂期,其核膜仍然存在,並不消失。細胞核與細胞質之間的界膜,厚約200埃,是真核細胞的特有結構。它除包括由雙層膜組成的“核膜”外,還包括兩層膜之間的核周腔和分佈在膜上的核孔。
內核膜包圍核質,並被核層覆蓋,能通過核孔複合體與外核膜相連。核層是由中間絲網組成的,能起到穩定核膜的作用,參與染色質功能和整個基因表達的過程。雖然內外核膜和內質網相連,但膜中嵌入的蛋白質傾向於保持在原有的區域上,而不是分散在整個連續體中,提示膜上可能還是有不連續的分界線。
內核膜蛋白的突變可引起幾種核包膜病,包括核纖層蛋白病等,部分癌症可能也和該位點病變相關。
核膜上有成千上萬個核孔,每個大的核孔複合體都有約100nm左右長,而在內核膜上的孔道大約有40nm寬,核孔能連接內外核膜。
在細胞間期的G2期,核膜表面積增加,核孔複合體數量增加一倍。在真核生物中,如酵母,在細胞分裂過程中,核膜保持完整。紡錘體纖維要麼在膜內形成,要麼穿透膜但不將其撕裂。在其他真核生物(動物和植物)中,核膜必須在有絲分裂的前期階段分解,使有絲分裂紡錘體纖維能夠進入其中的染色體。裂解和重建的具體機制還不完全了解清楚。
在哺乳動物中,核膜可以在幾分鐘內分解,而這在有絲分裂的早期階段遵循一系列步驟。首先,M-Cdk的磷酸化核蛋白多肽和核膜被選擇性地從核孔複合物中運出。之後,其餘的核孔複合物同時斷裂。不過現有生化證據表明核孔複合物分解成穩定的碎片,而不是分解成小的多肽碎片。M-Cdk還磷酸化了核層中的中間絲(支撐包膜的骨架),導致內層的分解。電子顯微鏡和熒光顯微鏡有力地證明了核膜被內質網-核蛋白所吸收,而在有絲分裂過程中,內質網中通常沒有發現內質網-核蛋白。
另外,除了在有絲分裂的前期階段核膜破裂之外,在細胞周期的間期階段,發生細胞遷移的哺乳動物細胞中,核膜也出現破裂。這種短暫的破裂可能由核變形引起。細胞質蛋白複合物組成的“運輸所需的內體分選複合物”(ESCRT)能快速修復破裂。在核膜破裂過程中,可能發生DNA雙鏈斷裂。因此,在受限環境中遷移的細胞的存活似乎依賴於有效的核包膜和DNA修復機制。
在核纖層蛋白病變的細胞和癌細胞中也觀察到異常的核膜破裂,這常常導致細胞蛋白質的錯位、微核的形成和基因組的不穩定性。
在細胞周期中解聚的核膜的重建具體機制尚不清楚,但主要有以下兩種假說。
1.原核膜解聚成為的囊泡重新聚合生成新的核膜。
2.新的內質網吸收原本核膜解聚生成的囊泡,而後包被原本的核區,形成封閉膜,進而生成核膜。
核膜的特殊作用就是把核物質集中在靠近細胞中央的一個區域內,核物質的區域化有利於實現其功能。
核膜對核內外物質的交通有高度選擇性,離子和小分子的通透是由核膜調節的,而核酸、蛋白質大分子的通透則是由核孔複合體的選擇通透性控制的。如果把原來存在於核內的蛋白質(如組蛋白、RNA 聚合酶等)注射到細胞質內,便能濃集到核內;而非核內蛋白,分子量超過 60000就很難通過核膜進入核內。推測核內蛋白質分子結構上可能含有特別的信號肽段,使它們能選擇地通過核膜而集中到核內。至於一些體積顯然大於核孔複合體有效通路的蛋白質和核蛋白顆粒(如核糖體亞基的分子大小達15納米)則假定是由於分子構象從球形變成棒形而得以通過的;或者由於與核孔邊沿的某些受體分子的相互作用,使孔徑擴大而得以通過的。
根據對核膜比較基因組學、進化、起源的研究,有科學家提出了原始真核生物“前核生物”(prekaryote)假說,認為其與古菌內共生最終觸發了核膜產生。
至於核膜的適應性功能,認為其可能是作為一種必要的屏障,保護基因組免受細胞線粒體前體產生的活性氧(ROS)的攻擊。
基本信息
- 中文名
- 核膜
- 外文名
- nuclear envelope
- 拼音
- hé mó
- 定義
- 位於真核生物的核與細胞質交界處的雙層結構膜
- 作用
- 控制細胞核內外物質和信息傳輸
- 膜層
- 雙層膜
- 分佈
- 真核生物的核與細胞質交界處
- 別名
- nucleolemma; karyotheca