轉錄
轉錄
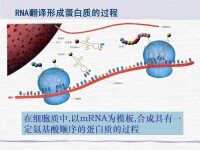
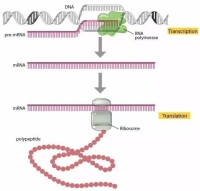
轉錄(Transcription)是遺傳信息從DNA流向RNA的過程。即以雙鏈DNA中的確定的一條鏈(模板鏈用於轉錄,編碼鏈不用於轉錄)為模板,以A,U,C,G四種核糖核苷酸為原料,在RNA聚合酶催化下合成RNA的過程。作為蛋白質生物合成的第一步,進行轉錄時,一個基因會被讀取並被複製為mRNA,即特定的DNA片斷作為遺傳信息模板,以依賴DNA的RNA聚合酶作為催化劑,通過鹼基互補的原則合成前體mRNA。RNA聚合酶通過與一系列組分構成動態複合體,完成轉錄起始、延伸、終止等過程。生成的mRNA攜有的密碼子,進入核糖體后可以實現蛋白質的合成。轉錄僅以DNA的一條鏈作為模板,被選為模板的單鏈稱為模板鏈,亦稱無義鏈;另一條單鏈稱為非模板鏈,即編碼鏈,因編碼鏈與轉錄生成的RNA序列T變為U外其他序列一致,所以又稱有義鏈。DNA上的轉錄區域稱為轉錄單位。
轉錄( transcription)是指以DNA為模板,以ATP、UTP、GTP和CTP為原料,按照鹼基互補原則,在RNA聚合酶的作用下合成RNA的過程,是基因c表達的第一步。原核細胞只有一種RNA聚合酶;而真核細胞則有3種:RNA聚合酶I、RNA聚合酶Ⅱ及RNA聚合酶Ⅲ。DNA雙鏈中作為轉錄模板的單鏈稱為模板鏈( templatestrand)或反義鏈( antisense strand),另一條鏈則稱為編碼鏈( coding strand)或有義鏈( sense strand)(圖2-6)。一個DNA分子上有許多基因,並非所有基因的編碼區都在同一條單鏈上,因此模板鏈或編碼鏈是相對某個基因的轉錄而言的。原核細胞的RNA轉錄:原核細胞的RNA聚合酶全酶(a2Bβ'σ)是由4條多肽鏈組成的核心酶加σ因子構成轉錄過程可劃分為開始、延伸和終止三個階段。①開始:σ因子識別DNA分子上的啟動子並與之結合,將DNA雙鏈局部解開,RNA合成開始,σ因子與核心酶分離。②延伸:RNA聚合酶沿模板鏈向前移動,使RNA鏈不斷合成延長。③終止:原核細胞轉錄終止分依賴ρ因子和不依賴p因子兩類。由於原核細胞沒有核膜,且合成的RNA幾乎不需進行複雜的加工修飾,故原核細胞的轉錄和翻譯兩個過程幾乎可以同時進行 。
轉錄過程
轉錄的概念以DNA為模板合成RNA,從而將遺傳信息傳遞到RNA分子中的過程即為轉錄。轉錄是RNA生物合成的最主要方式。轉錄需要DNA作模板,ATP、GTP、CTP、UTP等4種三磷酸核苷作為原料,RNA聚合酶催化作為模板的DNA在某一基因節段內只有一條鏈有轉錄功能,這條鏈稱為有意義鏈;而另一條鏈無轉錄功能,稱為反意義鏈。在DNA雙鏈上,各基因的有意義鏈不一定是同一條鏈,轉錄的這種選擇性稱為不對稱轉錄 。
以DNA為模板的RNA聚合酶,也稱轉錄酶。原核生物的RNA聚合酶分子量很大,通常由5個亞基組成;σ,β,β′和兩個α亞基,可寫作α2ββ′σ。含有5個亞基的酶叫全酶,失去σ亞基的叫核心酶(α2ββ′)。核心酶也能催化RNA的合成,但沒有固定的起始點,也不能區分雙鏈DNA的信息鏈與非信息鏈。σ亞基能識別模板上的信息鏈和啟動子,因而保證轉錄能從固定的正確位置開始。β和β′亞基參與和DNA鏈的結合。
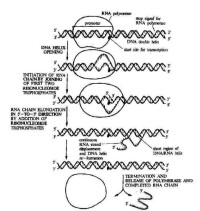
RNA聚合酶正確識別DNA模板上的啟動子並形成由酶、DNA和核苷三磷酸(NTP)構成的三元起始複合物(圖1),轉錄即自此開始。DNA模板上的啟動區域常含有TATAATG順序,稱普裡布諾(Pribnow)盒或P盒。複合物中的核苷三磷酸一般為GTP,少數為ATP(見核苷酸),因而原始轉錄產物的5′端通常為三磷酸鳥苷(pppG)或腺苷三磷酸(pppA)。真核 DNA上的轉錄啟動區域也有類似原核DNA的啟動區結構,和在-30bp(即在酶和 DNA結合點的上游30核苷酸處,常以-30表示,bp為鹼基對的簡寫)附近也含有TATA結構,稱霍格內斯(Hogness)盒或 TATA盒。第一個核苷三磷酸與第二個核苷三磷酸縮合生成3′-5′磷酸二酯鍵后,則啟動階段結束,進入延伸階段。
轉錄
轉錄的終止包括停止延伸及釋放 RNA聚合酶和合成的 RNA。在原核生物基因或操縱子的末端通常有一段終止序列即終止子; RNA合成就在這裡終止。原核細胞轉錄終止需要一種終止因子ρ(四個亞基構成的蛋白質)的幫助。
真核生物 DNA上也可能有轉錄終止的信號。已知真核DNA轉錄單元的3′端均含富有AT的序列〔如AATAA(A)或ATTAA(A)等〕,在相隔 0~30bp之後又出現TTTT順序(通常是3~5個T),這些結構可能與轉錄終止或者與3′端添加多聚A順序有關。
mRNA前體的后加工
原核mRNA的原始轉錄產物(除個別噬菌體外)都可直接用於翻譯,而真核mRNA一般都有相應的前體,前體必須經過後加工才能用於轉譯蛋白質。一般認為,真核mRNA的原始轉錄產物(也稱原始轉錄前體), hn RNA(hetero-geneous nuclear RNA,核不均一RNA),最終被加工成成熟的mRNA(見信使核糖核酸)。
mRNA前體的后加工包括以下四方面(圖2):①裝上 5′端帽子:轉錄產物的 5′端通常要裝上甲基化的帽子;有的轉錄產物 5′端有多餘的順序,則需切除后再裝上帽子。②裝上3′端多聚A尾巴:轉錄產物的3′端通常由多聚A聚合酶催化加上一段多聚A,多聚A尾巴的平均長度在20~200個核苷酸;有的轉錄產物的3′端有多餘順序,則需切除后再加上尾巴。裝5′端帽子和3′端尾巴均可能在剪接之前就已完成。③剪接:將mRNA前體上的居間順序切除,再將被隔開的蛋白質編碼區連接起來。剪接過程是由細胞核小分子RNA(如U1RNA)參與完成的,被切除的居間順序形成套索形(即lariat RNA中間體)。④修飾:mRNA分子內的某些部位常存在 N6 -甲基腺苷(見核苷酸),它是由甲基化酶催化產生的,也是在轉錄后加工時修飾的。

轉錄
目前分離得到的tRNA前體有兩類:①含單個tRNA的tRNA前體,在5′端和3′端各有一段多餘順序;②含二個tRNA的tRNA前體,除5′端和3′端有長短不一的多餘順序外,在兩個tRNA之間還有數目不等的核苷酸隔開。有的真核tRNA前體的反密碼子環區含有一個居間順序。
原核和真核生物tRNA前體的后加工有相似的步驟:①修飾:對tRNA分子上的部分核苷酸進行修飾(包括甲基化、醯化、硫代和重排等);②切除5′端和3′端多餘核苷酸;③3′端不含CCA順序的tRNA前體需裝上CCA順序(見轉移核糖核酸)。原核與真核tRNA前體的加工過程還有不同的情況:①原核多順反子tRNA前體,需加工時切開;②含有居間順序的真核tRNA前體,加工時需除去居間順序(圖3)。
轉錄
rRNA前體的后加工 通常有如下步驟:①修飾:除5SrRNA外,rRNA分子上通常有修飾核苷酸(主要是甲基化核苷酸),它們都是在後加工時修飾的。一般認為核糖2′羥基的甲基化在鹼基甲基化之前;②剪切:在rRNA前體分子的多餘順序處切開,產生許多中間前體,然後再切除中間前體末端的多餘順序(圖4);③剪接:有的真核生物rRNA前體中存在有居間順序的,須加工時除去。1982年T.R.切赫發現,在四膜蟲(Tetrahymena)rRNA前體中,去除含有413個核苷酸的居間順序是由rRNA前體自身催化完成的(圖5)。在 5′-鳥苷酸的促進下經過自身催化作用將居間順序切除,居間順序前後的兩個部分再連接起來,產生成熟的rRNA(5′-UpU-3′)和一個環狀 RNA分子及一個15個核苷酸殘基的小片段。rRNA前體的自身催化作用表明 RNA具有類似於酶的活性。這一發現突破了生物高分子中只有蛋白質才有催化作用的觀念。同時對生物進化與生命起源等研究都將有重要的意義。
是基因表達調節控制中的一個重要環節。促進基因轉錄叫正調節,抑制基因轉錄叫負調節。
轉錄
對真核細胞基因轉錄的調節控制目前知道得很少。同種高等生物每個個體的各個體細胞都有全套相同的基因,只是由於在發育過程中基因表達的調節控制(包括轉錄的調節控制)不同,因而發育成各種不同的組織和器官。目前認為,動物(包括人)都含有癌基因,但有的致癌,有的則不致癌,這也可能是由於轉錄與翻譯的調控不同。另外,真核DNA中的結構基因只佔總量的10%左右,大部分 DNA順序都可能起調節控制作用。真核生物也有誘導酶和誘導蛋白質,如干擾素就是由病毒或雙鏈RNA等誘導產生的一種蛋白質。轉錄抑製劑 轉錄能被一些特異性的抑製劑抑制,有些抑製劑是治療某些疾病的藥物,有的則是研究轉錄機理的重要試劑。按照作用機理的不同,轉錄抑製劑分為兩大類。第一類抑製劑特異性地與 DNA鏈結合,抑制模板的活性,使轉錄不能進行。這類抑製劑同時抑制DNA複製,例如:放線菌素D、紡錘菌素、遠黴素、溴乙錠和黃麴黴素等。第二類抑製劑作用於RNA聚合酶,使RNA聚合酶的活性改變或喪失,從而抑制轉錄的進行。這類抑製劑只抑制轉錄,不影響複製,是研究轉錄機制和RNA聚合酶性質的重要工具,例如:利福平。曲張鏈黴素、利鏈黴素和α-鵝膏蕈鹼等。
轉錄時,細胞通過鹼基互補的原則來生成一條帶有互補鹼基的mRNA,通過它攜帶密碼子到核糖體中可以實現蛋白質的合成。與DNA的複製相比,轉錄有很多相同或相似之處,亦有其自己的特點。
轉錄中,一個基因會被讀取並複製為mRNA。就是說,以特定的DNA片斷作為模板,以DNA依賴的RNA合成酶作為催化劑,合成前體mRNA。
在體內,轉錄是基因表達的第一階段,並且是基因調節的主要階段。轉錄可產生DNA複製的引物,在反轉錄病毒感染中也起到重要作用。
轉錄僅以DNA的一條鏈作為模板。被選為模板的單鏈叫模板鏈,又稱信息鏈、無義鏈;另一條單鏈叫非模板鏈。DNA上的轉錄區域稱為轉錄單位(transcription unit)。
RNA聚合酶合成RNA時不需引物,但無校正功能。
DNA: 5'-ATCGAATCG-3' (將此為非模板鏈)
3'-TAGCTTAGC-5' (將此為模板鏈)
轉錄出的 mRNA: 5'-AUCGAAUCG-3'
以RNA鏈為模板,經逆轉錄酶(即依賴於RNA的DNA聚合酶)催化合成DNA鏈,叫做逆轉錄。這種機制在RNA腫瘤病毒中首先發現。
RNA聚合酶是以DNA為模板的RNA聚合酶,也稱轉錄酶。
原核生物的RNA聚合酶分子量很大,通常由5個亞基組成;兩個α亞基,β,β′和σ,可寫作α2ββ′σ。含有5個亞基的酶叫全酶,失去σ亞基的叫核心酶(α2ββ′)。核心酶也能催化RNA的合成,但沒有固定的起始點,也不能區分雙鏈DNA的信息鏈與非信息鏈。σ亞基能識別模板上的信息鏈和啟動子,因而保證轉錄能從固定的正確位置開始。β和β′亞基參與和DNA鏈的結合。
真核生物RNA聚合酶有3類(不包括真核細胞線粒體中類似原核的RNA聚合酶),由8~12條亞基組成,分子量高達80萬。初步的研究指出,它們也可能存在類似原核的σ亞基組分。
在轉錄過程中,DNA模板被轉錄方向是從3′端向5′端;RNA鏈的合成方向是從5′端向3′端。RNA的
合成一般分兩步,第一步合成原始轉錄產物(過程包括轉錄的啟動、延伸和終止);第二步轉錄產物的后加工,使無生物活性的原始轉錄產物轉變成有生物功能的成熟RNA。但原核生物mRNA的原始轉錄產物一般不需后加工就能直接作為翻譯蛋白質的模板。
啟動
RNA聚合酶正確識別DNA模板上的啟動子並形成由酶、DNA和核苷三磷酸(NTP)構成的三元起始複合物,轉錄即自此開始。DNA模板上的啟動區域常含有TATAATG順序,稱普裡布諾(Pribnow)盒或P盒。複合物中的核苷三磷酸一般為GTP,少數為ATP,因而原始轉錄產物的5′端通常為三磷酸鳥苷(pppG)或腺苷三磷酸(pppA)。真核DNA上的轉錄啟動區域也有類似原核DNA的啟動區結構,和在-30bp(即在酶和DNA結合點的上游30核苷酸處,常以—30表示,bp為鹼基對的簡寫)附近也含有TATA結構,稱霍格內斯(Hogness)盒或 TATA盒。第一個核苷三磷酸與第二個核苷三磷酸縮合生成3′-5′磷酸二酯鍵后,則啟動階段結束,進入延伸階段。
延伸
σ亞基脫離酶分子,留下的核心酶與DNA的結合變松,因而較容易繼續往前移動。核心酶無模板專一性,能轉錄模板上的任何順序,包括在轉錄后加工時待切除的居間順序。脫離核心酶的σ亞基還可與另外的核心酶結合,參與另一轉錄過程。隨著轉錄不斷延伸,DNA雙鏈順次地被打開,並接受新來的鹼基配對,合成新的磷酸二酯鍵后,核心酶向前移去,已使用過的模板重新關閉起來,恢復原來的雙鏈結構。一般合成的RNA鏈對DNA模板具有高度的忠實性。RNA合成的速度,原核為25~50個核苷酸/秒,真核為45~100個核苷酸/秒。
終止
轉錄的終止包括停止延伸及釋放RNA聚合酶和合成的RNA。在原核生物基因或操縱子的末端通常有一段終止序列即終止子;RNA合成就在這裡終止。原核細胞轉錄終止需要一種終止因子ρ(四個亞基構成的蛋白質)的幫助。真核生物DNA上也可能有轉錄終止的信號。已知真核DNA轉錄單元的3′端均含富有AT的序列〔如AATAA(A)或ATTAA(A)等〕,在相隔0~30bp之後又出現TTTT順序(通常是3~5個T),這些結構可能與轉錄終止或者與3′端添加多聚A順序有關。
mRNA前體的后加工
原核mRNA的原始轉錄產物(除個別噬菌體外)都可直接用於翻譯,而真核mRNA一般都有相應的前體,前體必須經過後加工才能用於轉譯蛋白質。一般認為,真核mRNA的原始轉錄產物(也稱原始轉錄前體), hn RNA(hetero-geneous nuclear RNA,核不均一RNA),最終被加工成成熟的mRNA。
mRNA前體的后加工包括以下四方面:①裝上5′端帽子:轉錄產物的5′端通常要裝上甲基化的帽子;有的轉錄產物5′端有多餘的順序,則需切除后再裝上帽子。②裝上3′端多聚A尾巴:轉錄產物的3′端通常由多聚A聚合酶催化加上一段多聚A,多聚A尾巴的平均長度在20~200個核苷酸;有的轉錄產物的3′端有多餘順序,則需切除后再加上尾巴。裝5′端帽子和3′端尾巴均可能在剪接之前就已完成。③剪接:將mRNA前體上的居間順序切除,再將被隔開的蛋白質編碼區連接起來。剪接過程是由細胞核小分子RNA(如U1RNA)參與完成的,被切除的居間順序形成套索形(即lariat RNA中間體)。④修飾:mRNA分子內的某些部位常存在N6-甲基腺苷,它是由甲基化酶催化產生的,也是在轉錄后加工時修飾的。
有的真核mRNA前體,由於后加工的不同可產生兩種或兩種以上的mRNA(如人的降血鈣素基因轉錄產物),因而能翻譯成兩種或兩種以上的多肽鏈。
tRNA前體的后加工
目前分離得到的tRNA前體有兩類:①含單個tRNA的tRNA前體,在5′端和3′端各有一段多餘順序;②含二個tRNA的tRNA前體,除5′端和3′端有長短不一的多餘順序外,在兩個tRNA之間還有數目不等的核苷酸隔開。有的真核tRNA前體的反密碼子環區含有一個居間順序。
原核和真核生物tRNA前體的后加工有相似的步驟:①修飾:對tRNA分子上的部分核苷酸進行修飾(包括甲基化、醯化、硫代和重排等);②切除5′端和3′端多餘核苷酸;③3′端不含CCA順序的tRNA前體需裝上CCA順序。原核與真核tRNA前體的加工過程還有不同的情況:①原核多順反子tRNA前體,需加工時切開;②含有居間順序的真核tRNA前體,加工時需除去居間順序。首先,tRNA前體被一內切核酸酶將居間順序切除,產生帶有 2′,3′-環磷酸的5′半分子和含有5′羥基的3′半分子;然後兩個半分子分別在2′,3′-環磷酸二酯酶和多核苷酸激酶作用下使5′半分子露出了羥基和2′磷酸基,使3′半分子帶上5′磷酸基,這兩個半分子再先後經過連接酶和磷酸單酯酶(去除2′磷酸基)的作用,最後生成成熟的tRNA。
rRNA前體的后加工
rRNA前體的后加工通常有如下步驟:①修飾:除5SrRNA外,rRNA分子上通常有修飾核苷酸(主要是甲基化核苷酸),它們都是在後加工時修飾的。一般認為核糖2′羥基的甲基化在鹼基甲基化之前;②剪切:在rRNA前體分子的多餘順序處切開,產生許多中間前體,然後再切除中間前體末端的多餘順序;③剪接:有的真核生物rRNA前體中存在有居間順序的,須加工時除去。1982年T.R.切赫發現,在四膜蟲(Tetrahymena)rRNA前體中,去除含有413個核苷酸的居間順序是由rRNA前體自身催化完成的。在 5′-鳥苷酸的促進下經過自身催化作用將居間順序切除,居間順序前後的兩個部分再連接起來,產生成熟的rRNA(5′-UpU-3′)和一個環狀RNA分子及一個15個核苷酸殘基的小片段。rRNA前體的自身催化作用表明 RNA具有類似於酶的活性。這一發現突破了生物高分子中只有蛋白質才有催化作用的觀念。同時對生物進化與生命起源等研究都將有重要的意義。
真核生物RNA的轉錄與原核生物RNA的轉錄過程在總體上基本相同,但是,其過程要複雜得多,主要有以下幾點不同:
⒈ 真核生物RNA的轉錄是在細胞核內進行的,而蛋白質的合成則是在細胞質內進行的。所以,RNA轉錄后首先必須從核內運輸到細胞質內,才能指導蛋白質的合成。
⒉ 真核生物一個mRNA分子一般只含有一個基因,原核生物的一個mRNA分子通常含有多個基因,而除少數較低等真核生物外,一個mRNA分子一般只含有一個基因,編碼一條多肽鏈。
⒊ 真核生物RNA聚合酶較多,在原核生物中只有一種RNA聚合酶,催化所有RNA的合成,而在真核生物中則有RNA聚合酶Ⅰ、RNA聚合酶Ⅱ和RNA聚合酶Ⅲ三種不同酶,分別催化不同種類型RNA的合成。三種RNA聚合酶都是由10個以上亞基組成的複合酶。RNA聚合酶Ⅰ存在於細胞核內,催化合成除5SrRNA以外的所有rRNA的合成;RNA聚合酶Ⅱ催化合成mRNA前體,即不均一核RNA(hnRNA)的合成;RNA聚合酶Ⅲ催化tRNA和小核RNA的合成。
⒋ 真核生物RNA聚合酶不能獨立轉錄RNA 。原核生物中RNA聚合酶可以直接起始轉錄合成RNA ,真核生物則不能。在真核生物中,三種RNA聚合酶都必須在蛋白質轉錄因子的協助下才能進行RNA的轉錄。另外,RNA聚合酶對轉錄啟動子的識別,也比原核生物更加複雜,如對RNA聚合酶Ⅱ來說,至少有三個DNA的保守序列與其轉錄的起始有關,第一個稱為TATA框(TATA box),具有共有序列TATAAAA,其位置在轉錄起始點的上游約為25個核苷酸處,它的作用可能與原核生物中的-10共有序列相似,與轉錄起始位置的確定有關。第二個共有序列稱為CCAAT框(CCAAT box),具有共有序列GGAACCTCT,位於轉錄起始位置上游約為50-500個核苷酸處。如果該序列缺失會極大地降低生物的活體轉錄水平。第三個區域一般稱為增強子(enhancer),其位置可以在轉錄起始位置的上游,也可以在下游或者在基因之內。它雖不直接與轉錄複合體結合,但可以顯著提高轉錄效率。
轉錄能被一些特異性的抑製劑抑制,有些抑製劑是治療某些疾病的藥物,有的則是研究轉錄機理的重要試劑。按照作用機理的不同,轉錄抑製劑分為兩大類。第一類抑製劑特異性地與DNA鏈結合,抑制模板的活性,使轉錄不能進行。這類抑製劑同時抑制DNA複製,例如:放線菌素D、紡錘菌素、遠黴素、溴乙錠和黃麴黴素等。第二類抑製劑作用於RNA聚合酶,使RNA聚合酶的活性改變或喪失,從而抑制轉錄的進行。這類抑製劑只抑制轉錄,不影響複製,是研究轉錄機制和RNA聚合酶性質的重要工具,例如:利福平等。
在真核細胞中,RNA聚合酶通常不能單獨發揮轉錄作用,而需要與其他轉錄因子共同協作,有些輔助功能的轉錄因子就是解旋酶。
以反式作用影響轉錄的因子可統稱為轉錄因子(transcription factors, TF)。RNA聚合酶是一種反式作用於轉錄的蛋白因子。與RNA聚合酶Ⅰ、Ⅱ、Ⅲ相應的轉錄因子分別稱為TFⅠ、TFⅡ、TFⅢ,對TFⅡ研究最多。表19-2列出真核基因轉錄需要基本的TFⅡ。?
以前認為與TATA盒結合的蛋白因子是TFⅡ-D,後來發現TFⅡ-D實際包括兩類成分:與TATA盒結合的蛋白是TBP(TATAbox binding protein),是唯一能識別TATA盒並與其結合的轉錄因子,是三種RNA聚合酶轉錄時都需要的;其他稱為TBP相關因子(TBP?associated factors TAF),至少包括8種能與TBP緊密結合的因子。轉錄前先是TFⅡ-D與TATA盒結合;繼而TFⅡ-B以其C端與TBP-DNA複合體結合,其N端則能與RNA聚合酶Ⅱ親和結合,接著由兩個亞基組成的TFⅡ-F加入裝配,TFⅡ-F能與RNA聚合酶形成複合體,還具有依賴於ATP供給能量的DNA解旋酶活性,能解開前方的DNA雙螺旋,在轉錄鏈延伸中起作用。這樣,啟動子序列就與TFⅡ-D、B、F及RNA聚合酶Ⅱ結合形成一個“最低限度”能有轉錄功能基礎的轉錄前起始複合物(pre?intitiation complex, PIC),能轉錄mRNA。TFⅡ-H是多亞基蛋白複合體,具有依賴於ATP供給能量的DNA解旋酶活性,在轉錄鏈延伸中發揮作用;TFⅡ-E是兩個亞基組成的四聚體,不直接與DNA結合而可能是與TFⅡ-B聯繫,能提高ATP酶的活性;TFⅡ-E和TFⅡ-H的加入就形成完整的轉錄複合體,能轉錄延伸生成長鏈RNA,TFⅡ-A能穩定TFⅡ-D與TATA盒的結合,提高轉錄效率,但不是轉錄複合體一定需要的。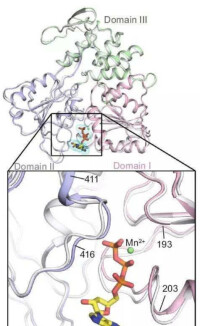
解旋酶

基本信息
- 中文名
- 轉錄
- 外文名
- Transcription
- 含義
- 遺傳信息由DNA轉換到RNA的過程
- 意義
- 基因調節的主要階段