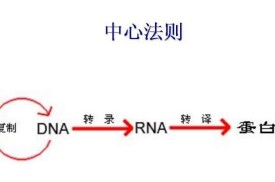
中心法則
遺傳信息在細胞內的生物大分子間轉移的基本法則
中心法則(英語:genetic central dogma),又譯成分子生物學的中心教條(英語:The central dogma of molecular biology) ,首先由佛朗西斯·克里克於1958年提出,並於1970年在《自然》上的一篇文章中重申:
徠“The central dogma of molecular biology deals with the detailed residue-by-residue transfer of sequential information. It states that such information cannot be transferred from protein to either protein or nucleic acid. (分子生物學的中心法則旨在詳細說明連串信息的逐字傳送。它指出遺傳信息不能由蛋白質轉移到蛋白質或核酸之中。)
是指遺傳信息從DNA傳遞給RNA,再從RNA傳遞給蛋白質,即完成遺傳信息的轉錄和翻譯的過程。也可以從DNA傳遞給DNA,即完成DNA的複製過程。這是所有有細胞結構的生物所遵循的法則。在某些病毒中的RNA自我複製(如煙草花葉病毒等)和在某些病毒中能以RNA為模板逆轉錄成DNA的過程(某些致癌病毒)是對中心法則的補充。
中心法則經常遭到誤解,尤其與遺傳信息“由DNA到RNA到“蛋白質””的標準流程相混淆。有些與標準流程不同的信息流被誤以為是中心法則的例外,其實朊病毒是中心法則現時已知的唯一例外。
遺傳信息的標準流程大致可以這樣描述:“DNA製造RNA,RNA製造蛋白質,蛋白質反過來協助前兩項流程,並協助DNA自我複製”.
遺傳信息在細胞內的生物大分子間轉移的基本法則。包含在脫氧核糖核酸(DNA)或核糖核酸(RNA)分子中的具有功能意義的核苷酸順序稱為遺傳信息。遺傳信息的轉移包括核酸分子間的轉移、核酸和蛋白質分子間的轉移。
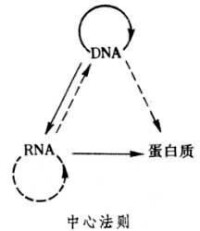
1957年F.H.C.克里克最初提出的中心法則是:DNA→RNA→蛋白質。它說明遺傳信息在不同的大分子之間的轉移都是單向的,不可逆的,只能從DNA到RNA(轉錄),從RNA到蛋白質(翻譯)。這兩種形式的信息轉移在所有生物的細胞中都得到了證實。1970年H.M.特明和D.巴爾的摩在一些RNA致癌病毒中發現它們在宿主細胞中的複製過程是先以病毒的RNA分子為模板合成一個DNA分子,再以DNA分子為模板合成新的病毒RNA。前一個步驟被稱為反向轉錄,是上述中心法則提出后的新的發現。因此克里克在1970年重申了中心法則的重要性,提出了更為完整的圖解形式。
這裡遺傳信息的轉移可以分為兩類:第一類用實線箭頭表示,包括DNA的複製、RNA的轉錄和蛋白質的翻譯,即①DNA→DNA(複製);②DNA→RNA(轉錄);③RNA→蛋白質(翻譯)。這三種遺傳信息的轉移方向普遍地存在於所有生物細胞中。第二類用虛線箭頭表示,是特殊情況下的遺傳信息轉移,包括RNA的複製,RNA反向轉錄為DNA和從DNA直接翻譯為蛋白質。即①RNA→RNA(複製);②RNA→DNA(反向轉錄);③DNA→蛋白質。RNA複製只在RNA病毒中存在。反向轉錄最初在RNA致癌病毒中發現,後來在人的白細胞和胎盤滋養層中也測出了與反向轉錄有關的反向轉錄酶的活性。至於遺傳信息從DNA到蛋白質的直接轉移僅在理論上具可能性,在活細胞中尚未發現。
克里克認為在圖解中沒有箭頭指向的信息轉移是不可能存在的,即①蛋白質→蛋白質;②蛋白質→RNA;③蛋白質→DNA。中心法則的中心論點是:遺傳信息一旦轉移到蛋白質分子之後,既不能從蛋白質分子轉移到蛋白質分子,也不能從蛋白質分子逆轉到核酸分子。克里克認為這是因為核酸和蛋白質的分子結構完全不同,在核酸分子之間的信息轉移通過沃森-克里克式的鹼基配對而實現。但從核酸到蛋白質的信息轉移則在現存生物細胞中都需要通過一個極為複雜的翻譯機構,這個機構是不能進行反向翻譯的。因此如果需要使遺傳信息從蛋白質向核酸轉移,那麼細胞中應有另一套反向翻譯機構,而這套機構在現存的細胞中是不存在的。中心法則合理地說明了在細胞的生命活動中兩類大分子的聯繫和分工:核酸的功能是儲存和轉移遺傳信息,指導和控制蛋白質的合成;而蛋白質的主要功能是進行新陳代謝活動和作為細胞結構的組成成分。
中子活化分析
以DNA為模板合成RNA是生物界RNA合成的主要方式,但有些生物像某些病毒,它們的遺傳信息貯存在RNA分子中,當它們進入宿細胞后,靠複製而傳代,它們在RNA指導的RNA聚合酶催化下合成RNA分子,當以RNA模板時,在RNA複製酶作用下,按5'→3'方向合成互補的RNA分子,但RNA複製酶中缺乏校正功能,因此RNA複製時錯誤率很高,這與反轉錄酶的特點相似。
RNA轉錄1
基因(DNA)主要位於細胞核中。如果酶(化學本質是蛋白質)是在細胞核內合成的,問題倒也簡單,由基因直接指導酶的合成就是了。可事實卻並不如此。
早在40年代,漢墨林(J·Hammerling)和布拉舍(J·Brachet)就分別發現傘藻和海膽卵細胞在除去細胞核之後,仍然能進行一段時間的蛋白質合成。這說明細胞質能進行蛋白質合成。1955年李托菲爾德(Littlefield)和1959年麥克奎化(K·McQuillen)分別用小鼠和大腸桿菌為材料證明細胞質中的核糖體是蛋白質合成的場所。這樣,細胞核內的DNA就必須通過一個“信使”(message)將遺傳信息傳遞到細胞質中去。
RNA轉錄2
實際上,早在1947年,法國科學家布瓦旺(A·Boivin)和旺德雷利(R·Vendrely)就在當年的《實驗》雜誌上聯名發表了一篇論文,討論DNA、RNA與蛋白質之間可能的信息傳遞關係。一位不知名的編輯把這篇論文的中心思想理解為DNA製造了RNA,再由RNA製造蛋白質。10年以後,1957年9月,克里克提交給實驗生物學會一篇題為“論蛋白質合成”的論文,發表在該學會的論文集(Symposum of the Society for Experimental Biology)第12卷第138頁。這篇論文被評價為“遺傳學領域最有啟發性、思想最解放的論著之一。”在這篇論文中,克里克正式提出遺傳信息流的傳遞方向是DNA→RNA→蛋白質,後來被學者們稱為“中心法則”。
生物遺傳中心法則最早是由Crick於1958年提出的,用以表示生命遺傳信息的流動方向或傳遞規律。由於當時對轉錄、翻譯、遺傳密碼、肽鏈摺疊等都還了解不多,在那個時候中心法則帶有一定的假設性質。隨著生物遺傳規律的進一步探索,中心法則也逐步得到完善和證實。
①1965年,科學家發現RNA可複製;
②1970年,科學家發現逆轉錄酶;
③1982年,科學家發現瘋牛病是由一種結構異常的蛋白質引起的疾病。
①從DNA流向DNA(DNA自我複製);
②從DNA流向RNA,進而流向蛋白質(轉錄和翻譯);
③從RNA流向RNA(RNA自我複製);
④從RNA流向DNA(逆轉錄)
註:其中前兩條是中心法則的主要體現,后兩條是中心法則的完善和補充。
中心法則是現代生物學中最重要最基本的規律之一,其在探索生命現象的本質及普遍規律方面起了巨大的作用,極大地推動了現代生物學的發展,是現代生物學的理論基石,並為生物學基礎理論的統一指明了方向,在生物科學發展過程中佔有重要地位。遺傳物質可以是DNA,也可以是RNA。細胞的遺傳物質都是DNA,只有一些病毒的遺傳物質是RNA。這種以RNA為遺傳物質的病毒稱為反轉錄病毒(retrovirus),在這種病毒的感染周期中,單鏈的RNA分子在反轉錄酶(reverse transcriptase)的作用下,可以反轉錄成單鏈的DNA,然後再以單鏈的DNA為模板生成雙鏈DNA。雙鏈DNA可以成為宿主細胞基因組的一部分,並同宿主細胞的基因組一起傳遞給子細胞。在反轉錄酶催化下,RNA分子產生與其序列互補的DNA分子,這種DNA分子稱為互補DNA(complementary DNA),簡寫為cDNA,這個過程即為逆轉錄(reverse transcription)。
由此可見,遺傳信息並不一定是從DNA單向地流向RNA,RNA攜帶的遺傳信息同樣也可以流向DNA。但是DNA和RNA中包含的遺傳信息只是單向地流向蛋白質,迄今為止還沒有發現蛋白質的信息逆向地流向核酸。這種遺傳信息的流向,就是克里克概括的中心法則(central dogma)的遺傳學意義。
任何一種假設都要經受科學事實的檢驗。反轉錄酶的發現,使中心法則對關於遺傳信息從DNA單向流入RNA做了修改,遺傳信息是可以在DNA與RNA之間相互流動的。那麼,對於DNA和RNA與蛋白質分子之間的信息流向是否只有核酸向蛋白質分子的單向流動,還是蛋白質分子的信息也可以流向核酸,中心法則仍然肯定前者。可是,病原體朊粒(Prion)的行為曾對中心法則提出了嚴重的挑戰。
基因指導蛋白質合成;基因控制生物體;生物體性狀由蛋白質直接體現。
a.基因通過控制酶的合成來控制代謝過程,進而控制生物體性狀;
b.基因通過指導蛋白質的合成,控制蛋白質結構進而直接控制生物體的性狀。
朊粒是一種蛋白質傳染顆粒(proteinaceous infectious particle),它最初被認識到是羊的瘙癢病的病原體。這是一種慢性神經系統疾病,在200多年前就已發現。1935年法國研究人員通過接種發現這種病可在羊群中傳染,意味著這種病原體是能在宿主動物體內自行複製的感染因子。朊粒同時又是人類的中樞神經系統退化性疾病如庫魯病(Kuru)和克—傑氏綜合征(Creutzfeldt-Jacobdisease,CJD)的病原體,也可引起瘋牛病即牛腦的海綿狀病變(bovin spongiform encephalopathy,BSE)。以後的研究證明,這種朊粒不是病毒,而是不含核酸的蛋白質顆粒。一個不含DNA或RNA的蛋白質分子能在受感染的宿主細胞內產生與自身相同的分子,且實現相同的生物學功能,即引起相同的疾病,這意味著這種蛋白質分子也是負載和傳遞遺傳信息的物質。這是從根本上動搖了遺傳學的基礎。
實驗證明,朊粒確實是不含DNA和RNA的蛋白質顆粒,但它不是傳遞遺傳信息的載體,也不能自我複製,而仍是由基因編碼產生的一種正常蛋白質的異構體。
哺乳動物細胞里的基因編碼產生一種糖蛋白PrP。人的PrP基因位於20號染色體短臂,PrP由253個氨基酸殘基組成,在氨基端有22個氨基酸組成的信號 肽。在正常腦組織中的PrP稱為PrPc,相對分子質量為33 000~35 000,對蛋白酶敏感。在病變腦組織中的PrP稱為PrPsc,相對分子質量為27 000~30 000,是PrPc中的一段,蛋白酶對其不起作用。現在知道,PrPc和PrPsc是PrP的兩種異構體,氨基酸組分和線性排列次序相同,但是三維構象不同。PrPc的結構中。螺旋佔42%,β片層佔30%;PrPsc則是。螺旋佔30%,β片層佔43%。PrPc的4條。螺旋可以排列成一個緻密的球狀結構,這個結構的隨機漲落(stochastic fluctuation)會長成部分摺疊的單體PrP*,這是一種中間體,即PrP*可以生成PrPc,也可以生成PrPsc。一般情況下,PrP*的含量極少,所以生成的PrPsc極少。可是外源的PrPsc可以促使PrP*變成PrPsc。PrPsc的不溶性使生成PrPsc過程成為不可逆轉。PrPsc在神經細胞里大量沉積,引起神經細胞的病變,破壞了神經細胞功能。因此,PrPsc感染正常細胞后,可以促使細胞內生成更多的PrPsc,PrPsc逐漸積累,需要有一個時間過程才會引發疾病,這也就是這種神經退化性疾病有一個很長的潛伏期的原因。所以說,PrPsc進入宿主細胞並不是自我複製,而是將細胞內基因編碼產生的PrPc變成PrPsc。由此可見,中心法則是正確的,至少在目前還是無需修正的。
中心法則是一個框架,用於理解遺傳信息在生物大分子之間傳遞的順序,對於生物體中三類主要生物大分子:DNA、RNA和蛋白質,有9種可能的傳遞順序。法則將這些順序分為三類,3個一般性的傳遞(通常發生在大多數細胞中),3個特殊傳遞(會發生,但只在一些特定條件下發生),3個未知傳遞(可能不會發生)。
| 法則中3類遺傳信息的傳遞順序 | ||
|---|---|---|
| 一般 | 特殊 | 未知 |
| DNA → DNA | RNA → DNA | 蛋白質→ DNA |
| DNA → RNA | RNA → RNA | 蛋白質→ RNA |
| RNA →蛋白質 | DNA →蛋白質 | 蛋白質→蛋白質 |
主條目:轉錄
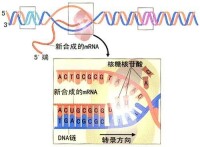
轉錄(Transcription)是遺傳信息由DNA轉換到RNA的過程。轉錄是信使RNA(mRNA)以及非編碼RNA(tRNA、rRNA等)的合成步驟。轉錄中,一個基因會被讀取、複製為mRNA;這個過程由RNA聚合酶(RNA polymerase)和轉錄因子(transcription factor)所共同完成。
主條目:RNA編輯
RNA編輯(RNA editing)是指在RNA水平上的改變遺傳信息的加工過程,導致成熟的RNA編碼序列和它的轉錄模板DNA序列之間的不相匹配。在真核生物的tRNA、rRNA和mRNA中都發現了RNA編輯這種現象。RNA編輯有核苷酸的刪除或插入編輯、鹼基替換編輯2種類型。這種改變影響了基因的表達,生成不同的氨基酸以及新的開放讀碼框。編輯可在多種水平被調節,並且與一些人類疾病有一定的相關性。
主條目:剪接 (遺傳學)
在真核細胞中,原始轉錄產物(mRNA前體)還要被加工:一個或多個序列(內含子)被剪出除去。選擇性剪接的機制使之可產生出不同的成熟的mRNA分子,這取決於哪段序列被當成內含子而哪段又作為存留下來的外顯子。並非全部有mRNA的活細胞都要經歷這種剪接;剪接在原核細胞中是不存在的。
主條目:翻譯 (遺傳學)
最終,成熟的mRNA接近核糖體,並在此處被翻譯。原核細胞沒有細胞核,其轉錄和翻譯可同時進行。而在真核細胞中,轉錄的場所和翻譯的場所通常是分開的(前者在細胞核,後者在細胞質),所以mRNA必須從細胞核轉移到細胞質,並在細胞質中與核糖體結合。核糖體會以三個密碼子來讀取mRNA上的信息,一般是從AUG開始,或是核糖體連接位下游的啟使甲硫氨酸密碼子開始。啟始因子及延長因子的複合物會將氨醯tRNA(tRNAs)帶入核糖體-mRNA複合物中,只要mRNA上的密碼子能與tRNA上的反密碼子配對,即可按照mRNA上的密碼序列加入氨基酸。當一個個氨基酸串連成胜肽鏈后,就會開始摺疊成正確的構形。這個摺疊的過程會一直進行,直到原先的多胜肽鏈從核糖體釋出,並形成成熟的蛋白質。在一些情況下,新合成的多胜肽鏈需要經過額外的處理才能成為成熟的蛋白質。正確的摺疊過程是相當複雜的,且可能需要其他稱為分子伴侶的幫忙。有時蛋白質本身會進一步被切割,此時內部被“捨棄”的部份即稱為內含肽。
主條目:DNA複製
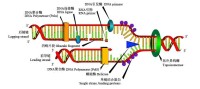
作為中心法則的最後一步,DNA必須忠實地進行複製才能使遺傳密碼從親代轉移至子代。複製是由一群複雜的蛋白質完成的;這些蛋白質打開超螺旋結構、DNA雙螺旋結構,並利用DNA聚合酶及其相關蛋白,拷貝或複製原模板,以使新代細胞或機體能重複DNA → RNA →蛋白質的循環。 DNA分子存在著構型多樣性,在遺傳信息的傳遞和表達過程中,DNA構象存在著左手螺旋及右手螺旋向右手螺旋的轉變過程,因此應賦有核酸構象的轉換形式。
有些病毒含有整套以RNA形式編碼的基因組,因此他們只有RNA→蛋白質的編譯形式。
主條目:擬逆轉錄
近年在植物體內發現了擬逆轉錄病毒(pararetrovirus),這種病毒的遺傳物質是雙鏈DNA,能像逆轉錄病毒一樣,通過把自己的DNA整合到寄主的基因組DNA中去,再進行複製。
主條目:逆轉錄
在中心法則被詳細闡述之後,人們發現了逆轉錄病毒。這些病毒可通過一種叫做逆轉錄酶的催化,以RNA為模板逆轉錄合成cDNA再由cDNA轉錄出RNA。這肯定了RNA向DNA轉錄的存在。人們最初以為這種現象僅出現於病毒中,但在最近,在高等動物中亦發現了RNA向DNA轉錄的逆轉錄轉座子。
主條目:RNA複製
有些病毒的遺傳物質是RNA分子,靠RNA複製而傳代,以RNA為模板的RNA複製酶催化下合成RNA分子,RNA複製酶中缺乏校正功能,複製時錯誤率很高。 RNA複製酶只對病毒本身的RNA起作用,而不會作用於宿主細胞中的RNA分子。
主條目:RNA催化
人們一直認為生物體內的各種生化反應都是由酶來催化完成的,而RNA僅是存貯與傳遞信息,與酶的催化反應無關。核糖核酸酶P是一種核酶,即由一個RNA分子發揮催化活性,它是第一個被發現的蛋白質以外具有催化活性的生物大分子。它的功能是剪切tRNA分子中RNA上多餘的或前體的多餘序列。RNA可以不通過蛋白質而直接表現出本身的某些遺傳信息,而這種信息並不是以核苷酸三聯密碼來編碼。
有人在一些離體實驗中觀察到,一些與蛋白質合成抑製劑類抗生素如新黴素和鏈黴素,能擾亂核糖體對信使的選擇,從而可以接受單鏈DNA分子代替mRNA,然後以單鏈DNA為模版,按核苷酸順序轉譯成多肽的氨基酸順序。另外還有研究表明,細胞核里的DNA可以直接轉移到細胞質中的核糖體上,不需要通過RNA也可以控制蛋白質的合成。
1994年喬依斯(G.F.Joyce)等人發現一個人工合成的DNA分子具有一種特殊的磷酸二酯酶活性。此後又有多例報道人工合成的DNA序列具有各種不同的酶活性。1995年中國學者王身立等人發現從多種生物中提取的DNA均具有酯酶活性,能催化乙酸萘酯水解為萘酚和乙酸。這種較弱的酯酶活性是非特異性DNA的一般性質,並不需要特定序列的DNA編碼。
克里克在上述那篇1970年的文章中指出,中心法則雖然對指導實驗很有用,但不應該被當成教條:
“雖然本文所提出的各類法則看來是可靠的,可是我們對分子生物學的認識,即使只是一個細胞—更不用說大自然里的整個生命體—仍然遠遠未完備到,足以讓我們把它當成教條一樣肯定正確的程度”
——克里克
自從克里克發表1970年那篇文章以來,很多新發現說明了中心法則補充和發展的必要。
主條目:翻譯后修飾
對於大部份的蛋白質來說,這是蛋白質生物合成的最後步驟。蛋白質的翻譯后修飾會附上其他的生物化學官能團、改變氨基酸的化學性質,或是造成結構的改變來擴闊蛋白質的功能。酶可以從蛋白質的N末端移除氨基酸,或從中間將肽鏈剪開。舉例來說,胰島素是肽的激素,它會在建立雙硫鍵后被剪開兩次,並在鏈的中間移走多肽前體,而形成的蛋白質包含了兩條以雙硫鍵連接的多肽鏈。其他修飾,就像磷酸化,是控制蛋白質活動機制的一部份。蛋白質活動可以是令酶活性化或鈍化。
主條目:蛋白質內含子
蛋白質有自剪接現象,與mRNA相同,一些蛋白質前體具有內含子(intein)序列,多肽序列中間的某些區域被加工切除,剩餘部分的蛋白質外顯子(extein)重新連接為蛋白質分子。
主條目:表觀遺傳學
表觀遺傳學研究在沒有細胞核DNA序列改變的情況時,基因功能的可逆的、可遺傳的改變。這些改變包括DNA的修飾(如甲基化修飾)、RNA干擾、組蛋白的各種修飾等。也指生物發育過程中包含的程序的研究。在這兩種情況下,研究的對象都包括在DNA序列中未包含的基因調控信息如何傳遞到(細胞或生物體的)下一代這個問題。其主要研究內容包括大致兩方面內容。一類為基因選擇性轉錄表達的調控,有DNA甲基化,基因印記,組蛋白共價修飾,染色質重塑。另一類為基因轉錄后的調控,包含基因組中非編碼的RNA,微小RNA,反義RNA,內含子及核糖開關等。
主條目:DNA甲基化
DNA甲基化為DNA化學修飾的一種形式,能在不改變DNA序列的前提下,改變遺傳表現。為外遺傳編碼(epigenetic code)的一部分,是一種外遺傳機制。DNA甲基化過程會使甲基添加到DNA分子上,例如在胞嘧啶環的5'碳上:這種5'方向的DNA甲基化方式可見於所有脊椎動物。
來自Mount.Sinai醫院的研究人員發現了一種叫Rev1 DNA聚合酶的蛋白質,它可以為DNA複製提供編碼信息。許多致癌物質會傾向於破壞DNA的鳥嘌呤(G),或者是破壞鳥嘌呤與胞嘧啶(C)的配對,這些都會導致DNA錯配的發生。新發現的蛋白質可以以自身為模板在複製鏈上加一個胞嘧啶,這個胞嘧啶無論鳥嘌呤是否在DNA鏈中存在都會被Rev1加上去的,在DNA複製時可以利用一條單鏈,根據鹼基配對原則複製出新的DNA鏈。細胞利用這種嶄新的機制在含有致癌物質的情況下對受損的DNA進行複製。這是第一次發現蛋白質可以作為一種合成DNA的模板。
主條目:朊病毒
朊病毒是通過改變其他蛋白質的構象來進行自身精確複製的一類蛋白質。也就是:蛋白質→蛋白質。這種具有感染性的因子主要由蛋白質組成。具有感染性的因子Prp與正常因子PrP在形狀上有一點不同。科學家推測這種變形的蛋白質會引起正常的PrP轉變成具有感染性的蛋白質,這種連鎖反應使得正常的蛋白質和致病的蛋白質因子都成為新病毒。
中心法則的信息是從DNA到RNA,但是,謝平(2014)指出,從生命起源和演化的歷史來看,信息的整合則必定是從mRNA到DNA 。
在徠細胞起源的早期,為了適應細胞的分裂行為,遺傳物質的有效傳遞成為必須,因此,細胞中儲存在各種m-RNA中的遺傳信息的整合必須成為選擇的方向,把所有m-RNA的信息連接起來,就是向DNA方向發展的啟航。也許可以認為,隨著蛋白質的增多,mRNA也相應增多,偶爾一個整合性的mRNA長鏈更好地匹配了細胞的分裂行為,這樣就會得到選擇。
但是,並不是把m-RNA拼接起來就是DNA,實際上,結構成份發生了兩個變化,其一,RNA分子中的尿嘧啶,在DNA中變成了胸腺嘧啶,雖然兩者僅有細微的差別,即後者多了一個甲基;其二,RNA分子中的核糖在DNA中變成了脫氧核糖。但是這兩個變化卻導致了兩種核酸在形態上的顯著差別:DNA形成雙螺旋的結構,而絕大部分RNA分子都是線狀單鏈,雖然RNA分子的某些區域可自身回折進行鹼基互補配對,形成局部雙螺旋。或許出於某種結構上的緣由,如果脫氧核糖替代核糖以及胸腺嘧啶替代尿嘧啶能更加有利於形成穩定的雙螺旋結構的話,那就是DNA被選擇的方向性。
當然,或許僅僅就是為了避免混淆,因為生物經常要用既有聯繫又能區別的結構物質來行使不同的功能,譬如,NADPH和NADH,兩者的還原電位完全相同,功能也類似,但卻用於不同的生物代謝途徑。一個不容易混淆的井然有序的代謝系統當然會得到選擇或青睞。
真核基因轉錄產物為單順反子,即一個基因編碼一條多肽鏈或RNA鏈,每個基因轉錄有各自的調節元件。在原核細胞中,通常是幾種不同的mRNA連在一起,相互之間由一段短的不編碼蛋白質的間隔序列所隔開,這種mRNA叫做多順反子mRNA。依筆者之見,原核生物多順反子的存在,正好可視為是mRNA拼接成長鏈DNA的一個過渡階段的證據。
DNA的複製
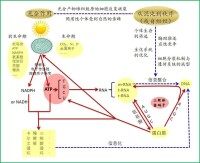
關於遺傳密碼系統起源的ATP中心假說
當然,完善的遺傳系統的建立絕非易事,超越了人類的想象,應該是細胞前體在數億年的演化歷程特別是無數次失敗的分裂過程中才得以實現的。人們可能懷疑這種推論的真實性,但在如此宏大的地球上,在如此之小的細胞中,如果給予了10億年的時日,一切偶然皆有可能成為必然,一切不可想象的事件皆有可能發生,只要有一個演化的方向性。
蛋白質、RNA和DNA三者在結構與功能的分化與完善,導致了一個完全獨立的遺傳系統的形成,而這又是通過細胞分裂維持生命形態相對穩定性的前提。只有一個真正可操作的遺傳系統的出現,生命才從前細胞體時代邁進了細胞時代,才真正拉開了生物進化的序幕。
在生化機制上,細胞必須形成既有區別又有聯繫的一種結構體系,即一方面必須對信息進行準確地儲存與複製,另一方面高效地實施生命構建,前者就是核酸體系,後者就是蛋白質體系。這兩個體系在短時間尺度上相對獨立,但不斷相互作用,導致在長時間尺度上的協同演化。
基本信息
- 中文名
- 中心法則
- 外文名
- genetic central dogma
- 提出時間
- 1958年
- 提出者
- 分類
- 醫學
- 作用
- 推動生物學發展
- 主要內容
- 完成遺傳信息的轉錄和翻譯的過程
- 又譯成
- 分子生物學的中心教條