磷酸二酯酶
水解細胞內第二信使
磷酸二酯酶磷酸二酯酶(PDEs)具有水解細胞內第二信使(cAMP,環磷酸腺苷或cGMP,環磷酸鳥苷)的功能,降解細胞內cAMP或cGMP,從而終結這些第二信使徠所傳導的生化作用。cAMP和cGMP對於細胞活動起著重要的調節作用。而其濃度的調節主要由核苷酸環化酶的合成和磷酸二酯酶(PDEs)水解作用之間的平衡決定。PDEs在人體內分佈廣泛,生理作用涉及多個研究領域。近年來,PDEs作為新的治療靶點,引起了眾多學者廣泛的關注,成為一個新的研究熱點,選擇性PDE4和PDE5抑製劑的臨床研究受到格外的重視。
分子克隆技術揭示磷酸二酯酶(phosphodiesterases,PDEs)是一個多基因大家族,開發選擇性的磷酸二酯酶抑製劑將為多種疾病的治療開闢新的思路。PDEs是一個多基因的大家族,它包括11型共30餘種具有不同底物專一性、酶動力學特徵、調控特點以及細胞與亞細胞分佈區域不同的磷酸二酯酶同功酶PDEs擁有相似的結構,均包含調控和催化兩個功能區域。各型PDEs催化區的氨基酸序列75%以上相同.顯示出家族成員間的同源性.並決定著對底物或抑製劑的專一性。PDEs具有不同的底物專一性:PDE4、7、8專一作用於cAMP,而PDE5、6、9則選擇性地作用於cGMP。PDE3以相似的親和力與cAMP和cGMP結合,但相對較不地水解cGMP,因而在功能上被視作對cAMP專一,cGMP通過與酶作用部位的競爭性結合而起負調控因子作用。PDEs1和.2既能水解cAMP,又能水解cGMP,但PDE1因其亞型不同,對兩種底物發揮不同的水解效能。PDEs的氨基端調控區域具有高度異源性,反映PDE家族成員的不同的輔助因子。此區域是與鈣調蛋白(CaM)(PDE1)、非催化的cGMP(PDE2、5、6)和轉導子(transducin)(PDE6)相結合的部位。另外,PDE3和PDE4氨基端部位還包括膜上的靶區域,PDE1、3、4、5包含蛋白激酶磷酸化部位。這些磷酸化部位能夠調節催化活性和(或)亞細胞定位 底物和輔助因子的恃異性組合使得cAMP和cGMP系統間的交互作用成為可能。在血小板,硝基擴管類藥物或PDE5抑製劑均可使cGMP增加,進而導致PDE3受抑制而繼發性地增加cAMP。相反,在腎上腺球粒細胞,心房利鈉因子(atrialⅡatri—tlret~factor,ANF)使cGMP增加並通過cGMP介導的PDE2活化,抑制cAMP刺激的醛固酮合成。
cAMP和cGMP作為神經遞質、激素、光和氣味等物質的第二信使,廣泛作用於細胞內靶器官,如:激酶、離子通道及各種PDEs。當外來信號經跨膜傳遞並引起一系列生理反應使核苷酸環化酶激活后(如圖1所示),cAMP和cGMP產生,PDEs家族的使命便是使之水解失活為5-單磷酸核苷(monophosphatenucleoside5,AMP)。核苷酸環化酶的合成和PDEs水解失活之間的平衡決定了第二信使cAMP和cGMP的濃度。值得注意的是,cGMP不但被PDEs水解,而且能調節一些PDEs活性,如PDE2可被cGMP刺激,而PDE3可被cGMP抑制,PDE4對cGMP不敏感。
PDE4抑製劑的抗炎作用機制主要涉及[5,8,9]:①抑制多種炎症介質/細胞因子的釋放,能夠抑制TH2細胞IL-4、IL-5基因的表達。②抑制白細胞的激活(呼吸爆發),抑制白細胞遊走。③抑制細胞粘附因子(CAM)的表達或上調。④誘導產生具有抑制活性的細胞因子,如IL-6。⑤誘導細胞凋亡。⑥刺激內源性激素和兒茶酚胺類物質的釋放。
PDE5作用機制 關於陰莖勃起的機制很複雜,目前尚無統一認識。現在多數研究者已將注意力從血流動力學轉向神經介質。已證實有三種神經機制共同參與海綿體平滑肌和螺旋動脈張力調控。即腎上腺素能、膽鹼能、和非腎上腺素能非膽鹼能(NANC)。現在認為NANC機制是海綿體血管、平滑肌舒張的主要機制,而NO為NANC的神經介質。在整個勃起生理過程中,陰莖血管及海綿體平滑肌舒張是關鍵因素。sildenafil通過NO/cGMP通路而發揮作用.在盆腔神經NANC神經系統刺激下,NO合酶(NOS)作用於L-精氨酸的胍基氮末端產生NO,由於其親脂性,可透過細胞膜在鄰近細胞間迅速擴散,並作用於細胞中的鳥苷酸環化酶(GC),與其亞鐵血紅素分子中的鐵離子結合,形成NO-GC複合物。這種複合物與該酶的卟啉部位相結合,導致構型的改變而激活GC,使細胞中cGMP大量產生,cGMP作為細胞內第二信使,可產生一系列生理反應,使海綿體平滑肌及血管舒張。cGMP傳遞信息的同時被PDE5水解轉化為5-GMP而失活。sildenafil選擇性抑制PDE5對cGMP水解,從而使cGMP積聚性濃度提高,大大增強了其血管舒張效果。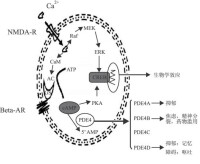
磷酸二酯酶
PDE1有3種同功酶:PDE1A、1B和1C,分別由不同的基因編碼。PDE1的催化活性是通過兩個CaM結合區域來調控的,然而每種同功酶都有其被激活的獨特Ca閾值。PDE1C可同等地水解cAMP和cGMP,能下調葡萄糖刺激的胰島素分泌。PDE1A和PDE1B主要水解cGMP。3種PDE1的分佈均有其確定的組織和細胞定位。PDE1B主要在腦和淋巴細胞表達,有絲分裂刺激后,其表達增加。目前已知的PDE1抑製劑有Vinpoeetin、Phenothiazine、SCH51866等,但它們缺乏特異性。鑒於PDE1基因的多樣性及其可能在Ca和核苷酸信號通路發揮交互聯繫的作用,提示其在治療中樞神經系統、心血管和免疫系統紊亂中有一定意義。
3種cGMP刺激的髓碴sPOE2A1、A2和A3都是一個基因的產物,但由於各外顯子連接的不同,它們的氨基酸存在差異。PDE2顯示出不同的組織和亞細胞分佈。膜結合的酶存在於腦和心臟.而可溶性的酶則存在於肝臟和血小板。PDE2亦分佈於T細胞.當抗原受體結合后,胸腺細胞PDE2活性下調。在胸腺細胞,依細胞內cGMP的濃度的不同.對cAMP代謝的控制可使PDE4轉換為PDE2。與此相似,在血小板,PDE2的作用有賴於環核苷酸的濃度:低cAMP濃度時,PDE2的活性依賴於cGMP;高cAMP濃度時,無論cGMP存在與否,PDE2水解cAMP。在心臟,cAMP和cGMP的濃度相互依賴,提示PDE2和PDE3抑製劑可在治療心絞痛、高血壓和心衰中發揮作用。EHNA[erythro-9一(2-hydroxy一3-nonytjadenine)為中等效能的PDE2選擇性抑製劑;也是腺苷脫氨酶的強抑製劑,因此可致腺苷聚集,並經由受體調控cAMP水平。cGMP和腺苷的協同作用也有利於心律失常的治療。
人類PDE3的兩種同功酶PDE3A和PDE3B是分別位於染色體12和11上的不同基因的產物。PDE3A和PDE3B的催化區域均包含一互不相同的44個氮基酸插入段。這44個氨基酸的不同不僅使PDE3A和PDE3B相互區分,亦使PDE3的催化部位有別於其它類型的PDEs。PDE3A和PDE3B的mRNA分佈於不同的組織和細胞:在血小板、心臟和血管平滑肌富含PDE3A,而PDE3B主要分佈在脂肪細胞和T淋巴細胞“許多PDE3抑製劑被視作強效血管擴張劑和兼有抗血小板活性的藥物而用於治療心衰,然而,前瞻性的隨機的Milrinone生存評估測試顯示:反覆口服給予PDE3抑製劑Milrlnone會增加心衰病人的死亡率。故僅少數PDE3抑製劑如Milrinone、Aminone、Enoximone可以急性、短期靜脈給葯治療心衰,同時須密切監測病人是否會出現室性心律失常增加的情況。PDE3B抑製劑可調節T淋巴細胞功能,人類T細胞以PDE3和PDE4佔優勢,PDE3和PDE4抑製劑相互協同,能強有力地抑制T細胞受體介導的細胞因子的產生和有絲分裂的擴增。
人類PDE4的同功酶具多樣性,分4種亞型:PDE4A、4B、4C和4D。PDE4與多種炎性細胞的cAMP水解有關,因此,抑制PDE4可抑制免疫和炎症細胞。而且,cAMP介導氣道平滑肌鬆弛。與口腎上腺素受體激動劑相比,PDE4抑製劑擴張支氣管的作用較弱。PDE4抑製劑Rolipram、Denbufylline、CDP840、CP80633、Ro201724等可能成為新型抗炎藥物。CDP840用於哮喘的治療.CP80633能減少特異反應性的病人的皮膚病變部位的炎症。選擇性PDE4亞型抑製劑的開發將有可能增強療效而減少不良反應。新型、強效PDE4抑製劑1-pyridy|naph—thalene的一系刊衍生物顯示出良好的抗哮喘作用且少有嘔吐及心血管方面的不良反應。
與PDE1和PDE2不同,PDE5絕對專一地水解cGMP,且無同功酶。PDE5包含兩個立體異構的cGMP結合位點,串聯地排列於蛋白質的氨基端cGMP與此兩位點的結合併不直接影響酶的催化活性,但影響酶被PKA磷酸化的能力。PDE5抑製劑有Zaprinast,ViagmCSilaenaft1),E4021等。cGMP中介NO介導的內皮鬆弛和ANF介導的利尿作用。通過抑制PDE5而使胞內cGMP升高將有利於高血壓、充血性心力衰竭、冠狀動脈疾病和心絞痛的治療,且PDE5抑製劑尚有拭血小板和抗血栓活性。此外,PDE5抑製劑能降低肺動脈壓力並對心率影響極少,有望成為新型的選擇性的肺動脈擴張劑。E-4010是一種選擇性的PDE5抑製劑,能夠緩解缺氧所致的大鼠肺動脈高壓。長期使用E-4010能降低monoer~aline所引發的肺動脈高壓大鼠的死亡率“FDA已批准Viagra用於治療男性陽痿和勃起功能障礙。人海綿體上主要分佈有PDE5,還有PDE2和PDE3l2。用PGE1和Forskolin研究結果提示陰莖平滑肌也可通過cAMP介導鬆弛。因此,Viagra尚有可能通過cGMP中介的PDE3抑制而升高cAMP發揮作用。在Viagra的Ⅰ期試驗中,有3的病人出現視覺徽染藍綠色,可能是由於視網膜功能的短暫的、可逆性的改變而造成。這主要是因為Viagra,E-4021等對PDE5和PDE6選擇性不高所造成。研製第三代PDE5抑製劑將對PDE5絕對專一性作用為方向。
PDE6是光感受器細胞進行光轉換級聯反應過程中重要的酶。它的活性受異三聚體G蛋白的調控。視桿細胞PDE6全酶是一個四聚體蛋白質,它包括α、β兩個大催化亞基和兩個具有PDE6抑制作用的小γ亞基。在暗反應光感受器,γ亞基有低水平的PDE6活性。遇光后,視色素視紫紅質激活視網膜G蛋白(即轉導子),使γ亞基發揮抑制PDE6的活性。而且,除了水解cGMP的活性部位,α、β兩個催化亞基有高親和力的非催化cGMP的結台位點,這些非催化位點不直接調控在活性位點的cGMP的催化,而是調控γ亞基對催化亞基的親和。
PDE7A1和PDE7A2是同一基因在表達上不同接合的變型,二者的mRNA均在多種組織中普遍表達。然而蛋白質的表達則有嚴格限制,提示PDE7的功能角色使其蛋白質的翻譯受到高度調控。在T淋巴細胞中已發現PDE7A1活性和蛋白質。抑制PDE7可能有利於治療某些免疫功能紊亂。
8其他磷酸二酯酶
目前徠,還認定了PDE8、PDE9、PDE10和PDE11。PDESA是一種cAMP選擇性酶,對非選擇性PDE抑製劑1BMX不敏感,但可被Dipyridimole所抑制。PDE9A是一種高度親和性的cGMP專一的PDE,可被Zaprinast輕度抑制。PDE9缺乏與PDE2、5、6相似的cGMP結合的立體異構區域。PDEgA的mRNA在腎臟高度表達Ⅲ。PDE10A水解cAMP和cGMP,其RNA在睾丸和腦組織中高度表達。
磷酸二酯酶超家族的多樣性和複雜性為多種疾病的治療提供了新的線索。必須設法了解PDEs作用的細胞內微環境及這些酶之間的相互影響和作用機制,對PDEs結構及PDEs抑製劑的研究有利於探索同功酶的選擇性,因而有利於新型高選擇性抑製劑的研製。
PDEs同工酶分佈在不同組織中,具有不同的生理功能。不只每個PDE家族有特異性底物和調節特點,而且每個家族各成員間也有組織、細胞、亞細胞特異性表達的不同,參與不同的信號傳導通路。選擇性PDEs抑製劑可特異性作用於不同的同工酶而發揮不同的作用。所以PDEs成為藥物開發很有吸引力的作用靶點。在心血管、生殖、抗炎、免疫等諸多領域均涉及到PDE同工酶家族。而且隨著分子生物學、生物化學、藥理學的研究發展,PDE同工酶被細分為若干亞家族及次亞家族,所以藥物可高特異性作用於某靶點,而大大減少毒副作用。對於選擇性PDEs抑製劑的研究,目前大多尚處於基礎試驗階段,臨床試驗剛剛起步。而且,新的家族成員不斷被發現,家族成員之間的差異也逐步達到能夠精細的定位。這預示藥理工作者在此領域將擁有一個艱辛而又前景無限的未來。

基本信息
- 中文名
- 磷酸二酯酶
- 外文名
- phosphodiesterases,PDEs
- 濃度的調節
- 傳導的生化作用
- 功能
- 水解細胞內第二信使