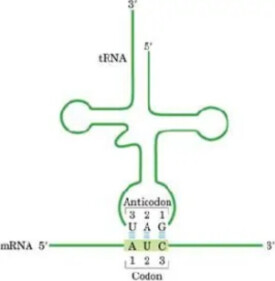
密碼子
三聯體密碼
密碼子(codon)是指信使RNA分子中每相鄰的三個核苷酸編成一組,在蛋白質合成時,代表某一種氨基酸的規律。
信使RNA在細胞中能決定蛋白質分子中的氨基酸種類和排列次序。信使RNA分子中的四種核苷酸(鹼基)的序列能決定蛋白質分子中的20種氨基酸的序列。而在信使RNA分子上的三個鹼基能決定一個氨基酸。
密碼子(codon):mRNA(或DNA)上的三聯體核苷酸殘基序列,該序列編碼著一個特定的氨基酸,tRNA 的反密碼子與mRNA的密碼子互補。
起始密碼子(iniation codon):指定蛋白質合成起始位點的密碼子。最常見的起始密碼子是甲硫氨酸或纈氨酸密碼。
終止密碼子(termination codon):任何tRNA分子都不能正常識別的,但可被特殊的蛋白質結合併引起新合成的肽鏈從翻譯機器上釋放的密碼子。存在三個終止密碼子:UAG,UAA和UGA。
可通過分析密碼子使用模式,預測目的基因的最佳宿主;或者應用基因工程手段,為目的基因表達提供最優的密碼子使用模式。3種不同的方式,目的都是利用密碼子偏愛性來提高異源基因的表達。
mRNA濃度是翻譯起始速率的主要影響因素之一,密碼子直接影響轉錄效率,決定mRNA濃度。如單子葉植物在“翻譯起始區”的密碼子偏性大於“翻譯終止區”,暗示“翻譯起始區”的密碼子使用對提高蛋白質翻譯的效率和精確性更為重要,因此,通過修飾編碼區5′端的DNA序列,來提高蛋白質的表達水平將有望成為可能。
基因的密碼子偏性與所編碼蛋白質結構域的連接區和二級結構單元的連接區有關、翻譯速率在連接區會降低。馬建民等通過聚類分析的方法研究發現,哺乳動物MHC基因的密碼子偏愛性與所編碼蛋白質的三級結構密切相關,並可通過影響mRNA不同區域的翻譯速度,來改變編碼蛋白質的空間構象。其研究所選取的蛋白質結構單位是蛋白指紋,它在很大程度上也是一種蛋白質功能單位,表明密碼子偏性與蛋白質的功能也存在密切相關。改變密碼子使用模式可目的性改變特定蛋白質的結構與功能。
密碼子的使用模式在細胞核和細胞質遺傳物質之間也存在差異,如核基因中的起始密碼子只有ATG,而線粒體基因中的起始密碼子為ATN;核基因中的終止密碼子TGA在線粒體基因中用來編碼色氨酸等。因此,可以通過比較密碼子的使用模式,來進行真核生物核糖體在細胞內以及未知基因在基因組的定位。
類似的密碼子使用模式,預示著物種相近的親緣關係或生存環境。目 前已有研究通過比較密碼子偏性的差異程度,來分析物種間的親緣關係和進化歷程。線粒體基因組具有母系遺傳、分子結構簡單、多態性豐富等優點,是一種重要的分子標記,研究其密碼子使用偏好性,可以很好地用於確定動物類群的遺傳分化和系統發生關係。

密碼子
遺傳信息是指DNA分子中基因上的脫氧核苷(鹼基)排列順序,密碼子是指信使RNA上決定一個氨基酸的三個相鄰鹼基的排列順序,反密碼子是指轉運RNA上的一端的三個鹼基排列順序。其聯繫是:DNA(基因)的遺傳信息通過轉錄傳遞到信使RNA上,轉運RNA一端攜帶氨基酸,另一端反密碼子與信使RNA上的密碼子(鹼基)配對
①. 遺傳密碼子是三聯體密碼:一個密碼子由信使核糖核酸(mRNA)上相鄰的三個鹼基組成。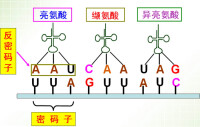 ② 密碼子具有通用性:不同的生物密碼子基本相同,即共用一套密碼子。
② 密碼子具有通用性:不同的生物密碼子基本相同,即共用一套密碼子。
密碼子
③ 遺傳密碼子無逗號:兩個密碼子間沒有標點符號,密碼子與密碼子之間沒有任何不編碼的核苷酸,讀碼必須按照一定的讀碼框架,從正確的起點開始,一個不漏地一直讀到終止信號。
④ 遺傳密碼子不重疊,在多核苷酸鏈上任何兩個相鄰的密碼子不共用任何核苷酸。
⑤ 密碼子具有簡併性:除了甲硫氨酸和色氨酸外,每一個氨基酸都至少有兩個密碼子。這樣可以在一定程度內,使氨基酸序列不會因為某一個鹼基被意外替換而導致氨基酸錯誤。
⑥ 密碼子閱讀與翻譯具有一定的方向性:從5'端到3'端。
⑦有起始密碼子和終止密碼子,起始密碼子有兩種,一種是甲硫氨酸(AUG),一種是纈氨酸(GUG),而終止密碼子(有3個,分別是UAA、UAG、UGA)沒有相應的轉運核糖核酸(tRNA)存在,只供釋放因子識別來實現翻譯的終止。
除了少數的不同之外,地球上已知生物的遺傳密碼均非常接近;因此根據演化論,遺傳密碼應在生命歷史中很早期就出現。現有的證據表明遺傳密碼的設定並非是隨機的結果,對此有以下的可能解釋:
最近一項研究顯示,一些氨基酸與它們相對應的密碼子有選擇性的化學結合力,這顯示現在複雜的蛋白質製造過程可能並非一早存在,最初的蛋白質可能是直接在核酸上形成。
尼倫伯格(M.W.Nirenberg,1927—)和馬太(H.Matthaei)破譯出了第一個遺傳密碼。
尼倫伯格和馬太採用了蛋白質的體外合成技術。他們在每個試管中分別加入一種氨基酸,再加入除去了DNA和mRNA的細胞提取液,以及人工合成的RNA多聚尿嘧啶核苷酸,結果加入了苯丙氨酸的試管中出現了多聚苯丙氨酸的肽鏈。實驗結果說明,多聚尿嘧啶核苷酸導致了多聚苯丙氨酸的合成,而多聚尿嘧啶核苷酸的鹼基序列是由許多個尿嘧啶組成的(UUUUUUUU......),可見尿嘧啶的鹼基序列編碼由苯丙氨酸組成的肽鏈。結合克里克得出的3個鹼基決定1個氨基酸的實驗結論,與苯丙氨酸對應的密碼子應該是UUU。在此後的六七年裡,科學家沿著蛋白質體外合成的思路,不斷地改進實驗方法,破譯出了全部的密碼子,並編輯出了密碼子表。
作用密碼錶
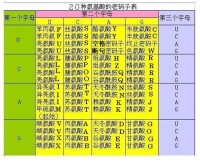
密碼子
依次根據氨基酸三字母縮寫,中文譯名拼音首字母尋找相關,再以其中密碼子簡併性(即重複性)最強的氨基酸為首選進行替代,具體變換為:
GCA,GCG: A→B
AGA,AGG: R→J
CCA,CCG: P→O
UUA,UUG: L→U
GUA,GUG: V→X
CAC : H→Z
對終止密碼子亦進行了調整。
需要強調,這一編碼方案忽略了既有的B、Z的使用,以及忽略了終止密碼子間真實的強弱性。
正向翻譯備選方案
A: GCU, GCC.
B: GCA, GCG.
C: UGU, UGC.
D: GAU, GAC.
E: GAA, GAG.
F: UUU, UUC.
G: GGU, GGC, GGA, GGG.
H: CAU.
I: AUU, AUC, AUA.
J: AGA, AGG.
K: AAA, AAG.
L: CUU, CUC, CUA, CUG.
M: AUG.
N: AAU, AAC.
O: CCA, CCG.
P: CCU, CCC.
Q: CAA, CAG.
R: CGU, CGC, CGA, CGG.
S: UCU, UCC, UCA, UCG, AGU, AGC.
T: ACU, ACC, ACA, ACG.
U: UUA, UUG.
V: GUU, GUC.
W: UGG.
X: GUA, GUG.
Y: UAU, UAC.
Z: CAC.
起始符:AUG【與M的編碼相同,但之後需加空格□(UAA)】
空格□:UAA
斷句符:UAG
終止符:UGA反向翻譯:見圖“修改後的密碼錶”
立體化學作用理論的代表人物是伍斯。他認為密碼起源於氨基酸和密碼子或反密碼子(或更一般地和RNA)的立體化學相互作用。這個觀點可以追溯至1962年,伍斯推測編碼關係可能是核酸與氨基酸間的立體化學作用,他把“簡併性”中涉及的密碼子看作是相等的核苷酸,1965年5月,伍斯發表題為《密碼的規則》的論文闡明遺傳密碼的排布規則,認為利用氨基酸色譜可以為分析“簡併性”提供有用的證據,傾向於密碼關係是一種核苷酸與氨基酸間的立體化學作用。此時,普適密碼還沒有完全確立,伍斯研究的編碼關係還具有一定的推測性。事實上,全部密碼關係的最佳立體化學匹配從來也沒有被證明過。但是,氨基酸的疏水性和反密碼予3’二核苷的疏水性順序相同是已確認的事實,說明立體化學作用的因素確實重要地影響著氨基酸和反密碼子的識別。
凍結偶然性理論的代表人物是克里克。該理論認為密碼關係是進化過程中的偶然性被固定下來的結果,這種關係一旦建立便永久保留下來。密碼子與氨基酸的對應關係是在某個生命發生時段里被固定下來,並且很難被改變,克里克在這篇論文中討論的編碼關係來源於他在1966年冷泉港會議上呈現的密碼錶,這張表除了起始密碼與UGA不同外,與今 天公認的普適密碼是基本一致的。儘管這個假說一直受到來自密碼的適應性、歷史性和化學特性三方面論點的挑戰,但是,從密碼結構的角度,不難看出克里克當時的密碼研究已經具有較強的客觀性和前瞻性了。另一方面,從氨基酸的生物合成看,在一條合成路徑上的幾個氨基酸所用編碼的密碼子往往只差一個鹼基。看來,後期形成的氨基酸和早期出現的氨基酸的編碼存在關係,這種氨基酸和密碼字典的協同進化說明編碼關係並非純屬偶然。
共進化假說提出傳統的密碼是從原始的簡單密碼進化而來,密碼子的進化與氨基酸生物合成的進化是並列的。主要證據是這個原始的密碼可能是由64個密碼子通過高度簡併只編碼少量的氨基酸,而後的進化中,那些來自相關合成路徑的物理化學性質不同的氨基酸卻具有相似的密碼子,表明密碼子的進化與氨基酸生物合成具有密切相關性。王子暉認為後引入的氨基酸密碼可能是通過篡奪與它具有相近生物合成路徑氨基酸的密碼而得到的,共進化理論鑒定了8組成對的前體和產物。這個假說後來還由朱利奧(M.Di Giulio)加以發展。
艾根等在研究遺傳密碼起源時進行試驗:在試管里沒有任何酶和模板的參與下,僅僅依靠鋅離子的催化,將核苷酸單體聚合成寡核苷酸,並通過彼此互為模板的複製、擴增,最終在不同條件的繼代培養下,優選出不同的tRNA克隆,然後形成RNA分子的准種群。這個實驗被稱為“試管選擇性理論”,證明在無生命力作用的情況下,自然條件下完全可以形成啟動生命形成的核糖核酸。根據地球形成之後的物理化學環境,推測生物大分子形成的活躍期是在距今38億年至40億年左右。依據實驗獲得的啟示,研究者認為地球的早期條件會影響早期短序列RNA的產生及密碼進化。但這個理論並沒有對密碼錶結構本身給予足夠的關注。
原始的遺傳密碼可能比今 天簡單得多,隨著生命演化製造出新的氨基酸再被利用而令遺傳密碼變得複雜。雖然不少證據證明這觀點,但詳細的演化過程仍在探索之中。經過自然選擇,現時的遺傳密碼減低了突變造成的不良影響。即,遺傳密碼是由選擇(selection)、歷史(history)和化學(chemistry)三個因素在不同階段起作用的(綜合進化假說)。
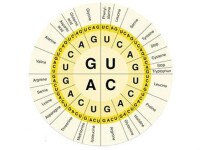
氨基酸
UUU (Phe/F)苯丙氨酸UUC (Phe/F)苯丙氨酸UUA (Leu/L)亮氨酸UUG (Leu/L)亮氨酸UCU (Ser/S)絲氨酸UCC (Ser/S)絲氨酸UCA (Ser/S)絲氨酸UCG (Ser/S)絲氨酸UAU (Tyr/Y)酪氨酸UAC (Tyr/Y)酪氨酸UAA (終止)UAG (終止)UGU (Cys/C)半胱氨酸UGC (Cys/C)半胱氨酸UGA (終止)UGG (Trp/W)色氨酸第 一 位 鹼 基 C
CUU (Leu/L)亮氨酸CUC (Leu/L)亮氨酸CUA (Leu/L)亮氨酸CUG (Leu/L)亮氨酸CCU (Pro/P)脯氨酸CCC (Pro/P)脯氨酸CCA (Pro/P)脯氨酸CCG (Pro/P)脯氨酸CAU (His/H)組氨酸CAC (His/H)組氨酸CAA (Gln/Q)谷氨醯胺CAG (Gln/Q)谷氨醯胺CGU (Arg/R)精氨酸CGC (Arg/R)精氨酸CGA (Arg/R)精氨酸CGG (Arg/R)精氨酸第 一 位 鹼 基 A
AUU (Ile/I)異亮氨酸AUC (Ile/I)異亮氨酸AUA (Ile/I)異亮氨酸AUG (Met/M)甲硫氨酸(起始)ACU (Thr/T)蘇氨酸ACC (Thr/T)蘇氨酸ACA (Thr/T)蘇氨酸ACG (Thr/T)蘇氨酸AAU (Asn/N)天冬醯胺AAC (Asn/N)天冬醯胺AAA (Lys/K)賴氨酸AAG (Lys/K)賴氨酸AGU (Ser/S)絲氨酸AGC (Ser/S)絲氨酸AGA (Arg/R)精氨酸AGG (Arg/R)精氨酸第 一 位 鹼 基 G
GUU (Val/V)纈氨酸GUC (Val/V)纈氨酸GUA (Val/V)纈氨酸GCU (Ala/A)丙氨酸GCC (Ala/A)丙氨酸GAU (Asp/D)天冬氨酸GAC (Asp/D)天冬氨酸GAA (Glu/E)谷氨酸GGU (Gly/G)甘氨酸GGC (Gly/G)甘氨酸GCA (Ala/ A)丙氨酸GCG (Ala/A)丙氨酸GUG (Val/V)纈氨酸GAG (Glu/E)谷氨酸GGG (Gly/G)甘氨酸語法及舉例語法a.以AUG表示進入正式信息編碼區;b.空格 (□) 用 UAA;c.逗號或其它文章內斷續標點用 UAG;d.編碼結束(最後一個句號)用 UGA表示;e. 默認為表達英語,如果需要表達其它語種,可以在起始信號(5'端)前延長6個鹼基,編碼語言使用語言的國家雙字母縮寫,如拼音加註CN,日語假名羅馬體加註JP等。舉例通用語言:I love U.中轉:(Start)□I□love□U.最終的核酸語言:AUG UAA AUU UAA CUG CCA GUC GAA UAA UUA UGA可以和連鶴或串珠等實體結構結合來進行空間變換,加強教學效果。但需要使遊戲者謹記這套密碼系統是修改過的。