ATP酶
生物膜上的蛋白酶之一
ATP酶又稱為三磷酸腺苷酶,是一類能將三磷酸腺苷(ATP)催化水解為二磷酸腺苷(ADP)和磷酸根離子的酶,這是一個釋放能量的反應。在大多數情況下,能量可以通過傳遞而被用於驅動另一個需要能量的化學反應。這一過程被所有已知的生命形式廣泛利用。
ATP是三磷酸腺苷的英文縮寫符號,它是各種活細胞內普遍存在的一種高能磷酸化合物。高能磷酸化合物是指水解時釋放的能量在20.92kJ/mol(千焦每摩爾)以上的磷酸化合物,ATP水解時釋放的能量高達30.54kJ/mol。ATP的分子式可以簡寫成A-P~P~P。簡式中的A代表腺苷,P代表磷酸基團,~代表一種特殊的化學鍵,叫做高能磷酸鍵。
ATP酶
部分ATP酶是內在膜蛋白,可以錨定在生物膜上,並可以在膜上移動;這些ATP酶又被稱為跨膜ATP酶。
ATP酶
ATP酶
2、構象假說;
3、化學滲透假說。
目前流行的是化學滲透假說,由英國生物化學家P.Mitchell於1961年提出。該學說很好地說明線粒體內膜中電子傳遞、質子電化學梯度建立、ADP磷酸化的關係,並具有大量的實驗支持,得到公認並獲得了1978年諾貝爾獎。
化學滲透假說的基本設想是:當高能電子沿呼吸鏈傳遞時,釋放出的能量使質子(H+)從線粒體內膜的基質側泵至膜間隙;內膜形成電化學質子梯度。在該梯度中蘊藏了能量,這種能量經ATP合成酶催化驅使ADP和無機磷酸形成ATP,即為氧化磷酸化過程。此假說依據線粒體的功能有四點具體的假設:
1、呼吸鏈各組成成分在線粒體內膜上有一定的位置。當電子從一種載體傳遞至另一種載體時,將質子泵出基質
2、線粒體ATP合成酶複合體也可跨膜轉運質子,但其作用是可逆的。該複合體利用足夠的電化學質子梯度能量在其內部合成ATP,這時質子由膜間隙通過複合體向基質方向流動;當電化學質子梯度不足以合成ATP時,ATP酶複合體能水解ATP,產生的能量將質子從基質側泵到膜間隙。
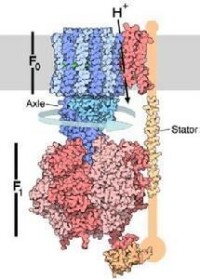
ATP酶
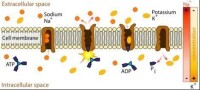
鈉鉀ATP酶的功能圖示
除了作為離子交換器,跨膜ATP酶還有其他類別,包括共轉運蛋白(co-transporter)和“泵”(也有部分“離子交換器”也被稱為“泵”)。這些跨膜ATP酶中,有一些可以造成膜內外電荷的流動,其他的則不行,因此又可以將這些轉運蛋白分為生電型(electrogenic)和非生電型。
人體預存的ATP能量只能維持15秒,跑完一百公尺后就全部用完,不足的繼續通過呼吸作用等合成ATP。純凈的ATP呈白色粉末狀,能溶於水,作為藥品可以提供能量並改善患者新陳代謝。ATP片劑可以口服,注射液可供肌肉注射或靜脈注射。
肌肉中儲藏著多種能源物質,主要有三磷酸腺苷(ATP)、磷酸肌酸(CP)、肌糖元、脂肪等。
(一)無氧代謝
劇烈運動時,體內處於暫時缺氧狀態,在缺氧狀態下體內能源物質的代謝過程,稱為無氧代謝。它包括以下兩個供能系統。
①非乳酸能(ATP—CP)系統—一般可維持10秒肌肉活動無氧代謝。
②乳酸能系統—一般可維持1—3分的肌肉活動非乳酸能(ATP—CP)系統和乳酸能系統是從事短時間、劇烈運動肌肉供能的主要方式。ATP釋放能量供肌肉收縮的時間僅為1—3秒,要靠CP分解提供能量,但肌肉中CP的含量也只能夠供ATP合成後分解的能量維持6—8秒肌肉收縮的時間。因此,進行10秒以內的快速活動主要靠ATP—CP系統供給肌肉收縮時的能量。乳酸能系統是持續進行劇烈運動時,肌肉內的肌糖元在缺氧狀態下進行酵解,經過一系列化學反應,最終在體內產生乳酸,同時釋放能量供肌肉收縮。這一代謝過程,可供1—3分左右肌肉收縮的時間。
(二)有氧代謝
在氧充足的條件下,肌糖元或脂肪徹底氧化分解,最終生成CO2和H2O,同時釋放大量的分解代謝,稱為有氧氧化系統。
(三)能量供應
1、了解體育促進身體健康的道理
體育運動加速體內能源物質的消耗,促進體內物質的分解與合成,使組織細胞得到比原有水平更多的營養補充,有機體獲得更加旺盛的活動能力,從而使身體不斷發展、完善,這就是體育鍛煉促進身體健康發展的基本道理。
2、了解能量供應與提高運動能力的關係
體育運動消耗體內的能源物質,經過一段時間休息后,體內能源物質可以恢復甚至超過原有水平,這種變化稱為超量恢復。出現超量恢復的程度和時間的早晚取決於運動量的大小。在一定範圍內運動量越大,體內能源物質消耗越多,超量恢復的幅度也越大,但所需的時間也長,在身體出現超量恢復階段,進行第二次適宜的運動與休息,可以逐步提高人體的能量供應水平,從而不斷提高人體運動能力。
3、了解有氧鍛煉與減肥的道理
長時間的運動是在有氧代謝的條件下進行的,要靠脂肪的代謝提供能量,因此,有氧運動是消耗脂肪達到減肥目的的有效方法。
4、人體的無氧代謝能力主要取決於以下三個方面:
①肌肉中ATP、CP的含量及分解速度;
②肌糖元的無氧酵解速度及血液對乳酸的緩衝能力;
③神經、肌肉對缺氧和乳酸堆積的耐受能力。
無氧代謝能力是速度素質的重要基礎。體育課發展無氧代謝能力的方法,一般採用間歇性練習和持續性練習。間歇練習主要發展ATP—CP系統的供能能力。一般每次練習在30秒以內,進行1—3分的積極性休息,再進行適宜練習,可以提高速度素質。持續練習主要發展乳酸系統的供能力。一般每次練習在30秒以上,每次休息時間較短,可以提高速度耐力。
5、發展有氧代謝能力
有氧代謝能力是人體長時間進行有氧運動的能力。發展有氧代謝能力關鍵在於有充足的氧供應,即人體單位時間內吸收、利用氧的最大數值——最大耗氧量。最大耗氧量與單位時間內血液循環攜帶、運輸氧有密切的關係。因此,心肺功能的好壞,直接影響到最大耗氧量。採用較低或中等運動強度、持續時間較長的練習,由於機體可以得到充足的氧供應,進行有氧氧化供能,所以,可以提高有氧代謝能力,從而提高心肺功能。
ATP作為一種輔酶,有改善肌體代謝的作用,可參與體內脂肪、蛋白質、糖、核酸、核苷酸等代謝過程。它同時又是體內能量的主要來源,為吸收、分泌、肌肉收縮以及進行生化合成反應等過程提供所需要的能量。常用於心肌病、肝炎、進行性肌萎縮、神經性耳聾等疾病的治療.
ATP廣泛用於改善機體代謝,以及疾病的輔助治療,是心臟病人常用的能量合劑中的重要成分之一。但心率過緩的病人要忌用,因為它會影響心室率和心臟的傳導,抑制心臟竇房結的正常工作,使其發出的衝動頻率減慢,傳導時間延長,導致心跳節律變慢。如果大劑量使用,可產生早搏、血壓下降等。因此,Ⅱ-Ⅲ度房室傳導阻滯、病態竇房結綜合征、交界性心律及洋地黃中毒引起高度房室傳導阻滯等疾病應忌用ATP,以免進一步減慢心律,心率低於60次/分應禁用ATP。
ATP除可引起上述副作用外,還可引起過敏性休克。近年來有關ATP引起過敏性休克甚至死亡的病例國內時有報道。其臨床表現為全身發抖、煩躁不安、呼吸困難和心律失常,因此使用時應注意如下幾個問題:
(1)應注意觀察有無過敏反應,凡過敏體質者不宜使用。
(2)ATP在體內分解后,能使全身血管擴張,血壓下降,因此它不宜應用於急性心肌梗塞。腦出血初期也應禁用ATP。ATP也不宜與能加重負性傳導和頻率作用的藥物合用。
(3)靜注時宜緩慢,應從小劑量開始治療,無效時可逐漸加量。
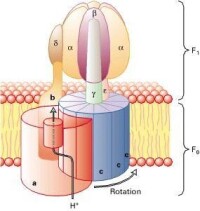
ATP合酶的結構
ATP合成酶可以在跨膜質子動力勢的推動下,利用ADP和Pi催化合成生物體的能量“通貨”——ATP。一般來說,機體所需的大多數ATP都是由ATP合酶產生的。據估計,人體每天進行正常活動所需的ATP量約等於他的體重,如體重70千克的成年人,每天合成的用於機體正常生命活動的ATP量約為70kg。而如此巨量的ATP正是由人體無數的ATP合酶合成的。
同時,ATP合成酶也可以催化逆反應,即ATP的水解。因此,從某種意義上來說,ATP合成酶也是一類ATP酶。
鈉/鉀離子轉運:ATP1A1, ATP1A2, ATP1A3, ATP1A4, ATP1B1, ATP1B2, ATP1B3, ATP1B4
鈣離子轉運:ATP2A1, ATP2A2, ATP2A3, ATP2B1, ATP2B2, ATP2B3, ATP2B4, ATP2C1
鎂離子轉運:ATP3
氫/鉀離子轉運:ATP4A, ATP4B
線粒體中的氫離子轉運:ATP5A1, ATP5B, ATP5C1, ATP5C2, ATP5D, ATP5E, ATP5F1, ATP5G1, ATP5G2, ATP5G3, ATP5H, ATP5I, ATP5J, ATP5J2, ATP5L, ATP5L2, ATP5O, ATP5S
溶酶體中的氫離子轉運:ATP6AP1, ATP6AP2, ATP6V1A, ATP6V1B1, ATP6V1B2, ATP6V1C1, ATP6V1C2, ATP6V1D, ATP6V1E1, ATP6V1E2, ATP6V1F, ATP6V1G1, ATP6V1G2, ATP6V1G3, ATP6V1H, ATP6V0A1, ATP6V0A2, ATP6V0A4, ATP6V0B, ATP6V0C, ATP6V0D1, ATP6V0D2, ATP6V0E
銅離子轉運:ATP7A, ATP7B
I型,第8類:ATP8A1, ATP8B1, ATP8B2, ATP8B3, ATP8B4
II型,第9類:ATP9A, ATP9B
V型,第10類:ATP10A, ATP10B, ATP10D
VI型,第11類:ATP11A, ATP11B, ATP11C
非胃中的氫/鉀離子轉運:ATP12A
第13類:ATP13A1, ATP13A2, ATP13A3, ATP13A4, ATP13A5
基本信息
- 中文名
- ATP酶
- 化學式
- A-P~P~P
- 類型
- 化合物
- 應用
- 被已知的生命形式廣泛利用
- 參與的反應
- 將三磷酸腺苷(ATP)催化水解為二磷酸腺苷(ADP)和磷酸根離子
- 全稱
- 三磷酸腺苷酶