鈉鉀泵
Na+-K+ATP酶
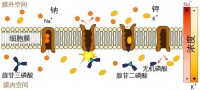
鈉鉀泵實際上就是Na+-K+ATP酶,一般認為是由2個大亞基、2個小亞基組成的4聚體。Na+-K+ATP酶通過磷酸化和去磷酸化過程發生構象的變化,導致與Na+、K+的親和力發生變化。在膜內側Na+與酶結合,激活ATP酶活性,使ATP分解,酶被磷酸化,構象發生變化,於是與Na+結合的部位轉向膜外側;這種磷酸化的酶對Na+的親和力低,對K+的親和力高,因而在膜外側釋放Na+、而與K+結合。
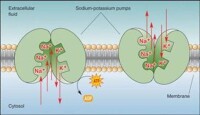
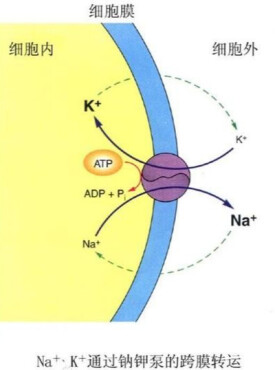
鈉鉀泵可以將細胞外相對細胞內較低濃度的鉀離子送進細胞,並將細胞內相對細胞外較低濃度的鈉離子送出細胞。經由以具放射性的鈉、鉀離子標定,可以發現鈉、鉀離子都會經過這個通道,鈉、鉀離子的濃度在細胞膜兩側也都是相互依賴的,所以顯示了鈉、鉀離子都可以經過這個載體運輸。且已知鈉鉀泵需消耗ATP,並可以將三個鈉離子送出細胞,同時將兩個鉀離子送進細胞。
鈉鉀泵在1950年被丹麥的科學家延斯·斯科(Jens Skou)發現,它代表了我們對離子進出細胞的認識的一個重要的里程碑。它也在細胞刺激上有著重要的意義,像神經細胞的衝動,就是用鈉鉀泵幫助維持細胞電位使神經衝動得以傳輸。
鈉鉀泵(也稱鈉鉀轉運體),為蛋白質分子,進行鈉離子和鉀離子之間的交換。每消耗一個ATP分子,逆電化學梯度泵出三個鈉離子和泵入兩個鉀離子。保持膜內高鉀膜外高鈉的不均勻離子分佈。
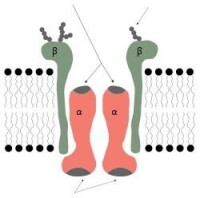
Na—K 泵由α、β兩亞基組成。α亞基為分子量約 120KD 的跨膜蛋白,既有Na、K 結合位點,又具 ATP 酶活性,因此 Na—K 泵又稱為 Na—K—ATP 酶。β
鈉鉀泵
一般認為 Na—K 泵首先在膜內側與細胞內的 Na 結合,ATP 酶活性被激活后,由 ATP 水解釋放的能量使“泵”本身構象改變,將Na 輸出細胞;與此同時, “泵”與細胞膜外側的 K 結合,發生去磷酸化后構象再次改變,將 K 輸入細胞內。研究表明,每消耗 1 個 ATP 分子,可使細胞內減少 3 個 Na 並增加 2 個 K。
細胞膜鈉鉀泵作用首先是由Hodkin和Keynes(1955)所發現.1957年Skou發現了Na+-K+ ATP酶並證明其與鈉鉀泵的作用有關.
鈉鉀泵的作用方式可因不同生理條件而異,在紅細胞膜中可能有以下幾種方式:
1.正常的作用方式——利用ATP的水介與Na+-K+的跨膜轉運相偶聯.
2.泵的反方向作用——利用Na+-K+的跨膜轉運來推動ATP的合成.
3.Na+ - Na+交換反應可能與ATP和ADP交換反應相偶聯.
4.K+ - K+交換反應與Pi和H2(18)O的交換反應相偶聯.
5.依賴ATP水解,解偶聯使Na+排出.
鈉鉀泵
鈉鉀泵的一個特性是他對離子的轉運循環依賴自磷酸化過程,ATP上的一個磷酸基團轉移到鈉鉀泵的一個天冬氨酸殘基上,導致構象的變化。通過自磷酸化來轉運離子的離子泵就叫做P-type,與之相類似的還有鈣泵和質子泵。它們組成了功能與結構相似的一個蛋白質家族 .Na-K泵作用是:①維持細胞的滲透性,保持細胞的體積;②維持低Na+高K+的細胞內環境,維持細胞的靜息電位.
靜息電位指安靜時存在於細胞兩側的外正內負的電位差。其形成原因是膜兩側離子分佈不平衡及膜對K+有較高的通透能力。細胞內K+濃度和帶負電的蛋白質濃度都大於細胞外(而細胞外Na+和Cl+濃度大於細胞內),但因為細胞膜只對K+有相對較高的通透性,K+順濃度差由細胞內移到細胞外,而膜內帶負電的蛋白質離子不能透出細胞,於是K+離子外移造成膜內變負而膜外變正。外正內負的狀態一方面可隨K+的外移而增加,另一方面,K+外移形成的外正內負將阻礙K+的外移(正負電荷互相吸引,而相同方向電荷則互相排斥)。最後達到一種K+外移(因濃度差) 和阻礙K+外移(正負電荷互相吸引,而相同方向電荷則相互排斥)。最後達到一種K+外移(因濃度差)和阻礙K+外移(因電位差)相平衡的狀態,這是的膜電位稱為K+平衡電位,實際上,就是(或接近於)安靜時細胞膜外的電位差。
能使Na+通道大量開放從而產生動作電位的臨界膜電位。(或能使膜出現Na+內流與去極化形成負反饋的膜電位值)稱為閾電位。在一定的刺激持續時間作用下,引起組織興奮所必需的最小刺激強度,稱為閾強度。比閾電位弱的刺激,成為閾下刺激,他們只能引起低於閾電位值的去極化,不能發展為動作電位。閾下刺激未能使靜息電位的去極化達到閾電位,但他也能引起該段膜中所含Na+通道的少量開放,這是少量Na+內流造成的去極化和電刺激造成的去極化疊加起來,在受刺激的局部出現一個較小的去極化,成為局部興奮或局部反應。其特點為:①它不是“全或無”的,在閾下刺激的範圍內,隨刺激強度的增大而增大,②不能在膜上作遠距離的傳播,但由於膜本身由於有電阻和電容特性而膜內外都是電解質溶液,發生在膜的某一點的局部興奮,可以使鄰近的膜也產生類似的去極化,但隨距離加大而迅速減小以至消失,成為電緊張性擴布③局部興奮可以互相疊加,當一處產生的局部興奮由於電緊張性擴布致使臨近處的膜也出現程度較小的去極化,而該處又因另一刺激也產生了局部興奮,雖然兩者單獨出現時都不足以引起一次動作電位,但如果遇到一起時可以疊加起來,以致有可能達到閾電位引發一次動作電位,稱為空間性總和。局部興奮的疊加也可以發生在連續數個閾下刺激的膜的某一點,亦即當前面刺激引起的局部興奮尚未消失時,與後面刺激引起的局部興奮發生疊加,稱為時間性總和。在刺激超過閾強度后,動作電位的上升速度和所能達到的最大值,就不再依賴於所給刺激的強度大小了。即只要刺激達到足夠的強度,再增加刺激強度並不能使動作電位的幅度有所增大。此外,動作電位並不是只出現在受刺激的局部,他在受刺激部位產生后,還可沿著細胞膜向周圍傳播,而且傳播的距離並不因為原處刺激的強度而有所不同,直至整個細胞的膜都依次興奮併產生一次同樣大小和形式的動作電位。即動作電位的“全或無”現象。
經科學研究,發現Na+-K+泵在人體的正常代謝中具有非常重要的作用,與一些疾病的發生也有著密切的關係。如肝水腫,白內障,囊纖維化 ,癲癇 ,偏頭痛 ,高血
鈉鉀泵
在這裡,僅就白內障和高血壓與Na+-K+泵的關係做一點介紹.
白內障的病因 較為複雜,可能是環境,營養,代謝和遺傳等多種因素。對晶狀體長期綜合作用的結果。一般認為,氧化損傷引起白內障的最早期變化。氧化作用會損傷晶狀體細胞膜,使維持細胞內正常代鈉和高鉀離子濃度的Na+-K-ATP酶泵功能明顯改變,對鈉離子的通透性增加,使晶狀體內的鈉離子增加,導致水的流失,開始了皮質性白內障的過程
據報道,高血壓患者及有高血壓家族史而血壓正常者有跨膜電解質轉運紊亂,其血清中有一種激素樣物質,可抑制Na+/K+-ATP酶活性,以致鈉鉀泵功能降低,導致細胞內Na+,Ca2+濃度增加,動脈壁SMC收縮加強,腎上腺素能受體(adrenergic receptor)密度增加,血管反應性加強。這些都有助於動脈血壓升高。近來研究發現,血管緊張素(AGT)基因可能有15種缺陷,正常血壓的人偶見缺陷,而高血壓患者在AGT基因上的3個特定部位均有相同的變異。患高血壓的兄弟或姐妹可獲得父母的AGT基因的同一拷貝。有這種遺傳缺隱的高血壓患者,其血漿血管緊張素原水平高於對照組.
新近發現,有些肥胖者雖然堅持節食,活動量也不小,但依然"體壯膘肥",即使使盡各種減肥手段,體重也有增無減,煞是令人苦惱。其實,這種肥胖的根本原因是因為人體中褐色脂肪組織的產熱功能發生了故障,無法正常產熱,不能消耗能源脂肪.這主要是鑲嵌在竭色脂肪細胞膜上的一種被稱為鈉鉀三磷酸腺苷酶的"轉移泵"運轉慢了,由於"泵機"轉運減速,以燃燒脂肪為主的產熱機器便無法正常運行,使人的基礎體溫降低,機體耗能也減少。這種人好像處於一種亞冬眠的低能耗狀態,能量消耗少,人也就瘦不了.
由於Na+-K+泵在人體的生命活動中具有如此重要作用,更與疾病有著千絲萬縷的關係,我們相信在不久的將來,隨著的研究的深入,人們一定能解決許許多多的未解之謎!
除 Na—K 泵外,還有與 Ca、H 轉運有關的 Ca 泵和質子泵。這些“泵”的作用,對於維持細胞內環境的穩定亦具有重要意義。
總結:人體吸鉀排鈉
基本信息
- 中文名
- 鈉鉀泵
- 外文名
- sodium potassium pump
- 別名
- 鈉鉀ATP酶
- 應用
- 防止細胞水腫;勢能貯備
- 原理
- 保持膜內高鉀膜外高鈉的分佈
- 生物現象
- 靜息電位及動作電位