操縱子學說
操縱子學說
徠操縱子學說是1961年,法國科學家莫諾(J·L·Monod,1910-1976)與雅可布(F·Jacob)發表“蛋白質合成中的遺傳調節機制”一文中提出的學說,該學說開創了基因調控的研究。
色氨酸操縱子
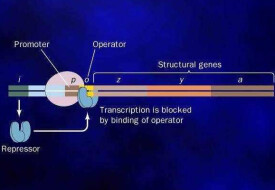
乳糖可作為培養大腸桿菌的能源。大腸桿菌能產生一種酶(叫做“半乳糖苷酶”),能夠催化乳糖分解為半乳糖和葡萄糖,以便作進一步的代謝利用。編碼半乳糖苷酶的基因(簡稱z)是一個結構基因(structural gene)。這個結構基因與操縱基因共同組成操縱子。操縱基因受一種叫作阻遏蛋白的蛋白質的調控。當阻遏蛋白結合到操縱基因之上時,乳糖會起誘導作用,它與阻遏蛋白結合,使之從操縱基因上脫落下來。這時,操縱基因開啟,相鄰的結構基因也表現活性,細菌就能分解並利用乳糖了,這樣,乳糖便成了誘導半乳糖苷酶產生的誘導物。
上述內容表明,大腸桿菌的乳糖操縱子是一個十分巧妙的自動控制系統:當培養基中含有充分的乳糖,同時不含葡萄糖時,細菌便會自動產生半乳糖苷酶來分解乳糖,以資利用。當培養基中不含乳糖時,細菌便自動關閉乳糖操縱子,以免浪費物質和能量。
60年代中期,在操縱子中還發現了另一個開關基因,稱為啟動基因(promoter)。啟動基因位於操縱基因之前,二者緊密相鄰。啟動基因由環腺苷酸(cAMP)啟動,而環腺苷酸能被葡萄糖所抑制。這樣,葡萄糖便通過抑制環腺苷酸而間接抑制啟動基因,使結構基因失活,停止合成半乳糖苷酶。
徠由此可知,結構基因同時受兩個開關基因——操縱基因與啟動基因的調控。只有當這兩個開關都處於開啟狀態時,結構基因才能活化。當培養基中同時存在葡萄糖和乳糖時,葡萄糖通過抑制環腺苷酸而間接抑制啟動基因,並進而抑制結構基因,使細菌不產生半乳糖苷酶。這種情況下,細菌便會自動優先利用葡萄糖,因為葡萄糖果是比乳糖更好的能源。
1969年,貝克維斯(J·R·Beckwith)從大腸桿菌的DNA中分離出乳糖操縱子,完全證實了雅可布和莫諾的模型。
在啟動基因發現之前,莫諾和雅可布的操縱子模型中,直接對結構基因起操縱作用的開關基因,僅有一個操縱基因。因此,有人開玩笑說:“半個操縱子就可以得諾貝爾獎”。對某一項成就,人們如果說它的一半就可以實現某種重要作用,就表明這項成就的偉大。北宋時代的名臣趙普就有“半部論語就可以治天下”的名言,由此也可見操縱子學說的巨大意義。
一個控制細胞基因表達的模型稱為操縱子(operon),此模型的提出使基因概念又向前邁出了一大步。表明人們已認識到基因的功能並不是固定不變的,而是可以根據環境的變化進行調節。隨之人們發現無論是真核還是原核生物轉錄調節都是涉及到編碼蛋白的基因和DNA上的元件。這一發現獲得了1965年諾貝爾獎。一個基因就是一段編碼有功能產物的DNA順序。基因的產物可以是蛋白質或是RNA(如tRNA和rRNA)。基因的重要特點是在有的情況下其產物能從合成位點散開去作用別的位點。
DNA元件是DNA上一段順序,它不能轉變成任何其它的形式,但它作為一種原位(in situ)順序具有特殊的功能。由於它只能作用同一條DNA,因此稱順式作用元件(cis-acting element)。
基因可以根據它們的產物分成不同的類型。編碼細胞必要的蛋白,如酶或結構蛋白的基因稱為結構基因( structural genes)。這類基因在細胞中占絕大部分,承擔著細胞各種蛋白的結構和功能。編碼調節蛋白的基因稱調節基因(regulator genes)。調節蛋白可調節其它基因的表達。由於調節基因的產物可以自由地結合到其相應的靶上,因此被為反式作用因子(trans-acting因子)。
調節的關鍵是調節基因編碼調節蛋白,此蛋白通過和DNA上特殊位點的結合來控制轉錄。順式作用元件被反式作用因子識別可以以“正”的或“負”的形式調節靶基因,順式作用位點通常總是在靶基因的上游。