腺病毒
無包膜的雙鏈DNA病毒
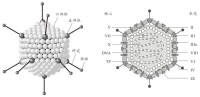
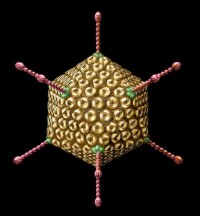
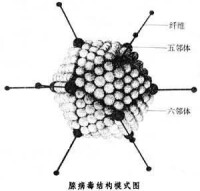
腺病毒(adenovirus)是一種沒有包膜的直徑為70~90 nm的顆粒,由252個殼粒呈廿面體排列構成。每個殼粒的直徑為7~9 nm。
腺病毒衣殼裡是線狀雙鏈DNA分子,約含4.7kb,兩端各有長約100 bp的反向重複序列。由於每條DNA鏈的5'端同相對分子質量為55X10Da的蛋白質分子共價結合,可以出現雙鏈DNA的環狀結構。
腺病毒
腺病毒對嚙齒類動物有致癌能力,或能轉化體外培養的嚙齒類動物細胞。使細胞轉化只需要腺病毒基因組的一部分,這些基因位於基因組的左端,約佔整個基因組的7%~10%。儘管腺病毒分佈很廣,但對人體不出現致癌性。人體細胞是一類允許細胞(permissive cell),即這類細胞允許感染入侵的病毒在細胞內複製增殖,最後細胞裂解死亡而釋放出大量子代病毒。在體外培養的多種人體腫瘤細胞中均未查出腺病毒顆粒,但在人的1號染色體上有adl2的整合位點,這意味著人體細胞對於腺病毒也可能是非允許細胞,即這類細胞在病毒感染后,病毒不能在細胞內複製增殖,但可整合在受感染細胞的基因組內。這些細胞被病毒轉化,表型發生改變,且可在體外無限期地培養傳代。
腺病毒結構示意圖
腺病毒含13%DNA和87%的蛋白質,病毒體分子量約為175×10 。病毒基因組為線狀雙鏈DNA,大約含35kb~36kb,腺病毒12、18和31型的DNA組成中,G+C mol%最低(48%~49%),屬於對動物具有高致癌性基因型。腺病毒1、2、4、5、8等型的G+C mol%較高(61%),致癌性反而低或無。這是一種用於人腺病毒分離株的分組的標準,根據其基因同源性將人腺病毒分為A~F等6組。
腺病毒的基因組以線性的雙鏈DNA形式存在,由蛋白VII和一種稱為mu的小蛋白緊密地環繞在其周圍,起到類組蛋白樣的作用。另一種蛋白V將這種DNA-蛋白複合物連接起來,並通過蛋白VI與病毒衣殼連接在一起。在兩條鏈的5′端各以共價鍵結合著一個被稱為DNA末端蛋白(pTP)複合物(DNA-TPC)的特化的結構,與腺病毒複製密切相關。腺病毒基因組的兩端各有一段100bp的反向末端重複序列(ITR),是複製的起始位點。在左端ITR的3′側有一段長約300bp的包裝信號(ψ)介導腺病毒基因組包裝入病毒衣殼。對腺病毒而言,只有包括兩端的ITR和包裝信號(ψ)的約0.5kb的序列是順式作用元件,也就是說必須由腺病毒載體自身攜帶,而其他的30餘種蛋白都可以通過輔助病毒(或細胞)反式補足。
病毒蛋白約11種(TP和PⅠ~PⅩ),其中有4種蛋白(病毒多肽PⅤ、PⅦ,末端蛋白TP、酶蛋白PⅩ)與病毒基因構成病毒核心,多肽PⅦ是主要的核心蛋白,如同組蛋白一樣包裹病毒基因DNA。構成病毒衣殼的蛋白質約7種。多肽PⅡ是病毒衣殼中最豐富和最主要成分,六鄰體是由3個PⅡ分子緊密相連組成。多肽PⅥ、PⅧ在六鄰體與病毒核心之間形成連接橋,並與多肽PⅨ一起穩定著六鄰體分子的晶格排列。5個分子多肽PⅢ相連構成五鄰體的基座蛋白,PⅢa為五鄰體的周圍蛋白,也參與衣殼的組成,五鄰體通過PⅤ與病毒核心相連。多肽PⅣ主要構成病毒三聚體纖突,纖突與病毒血凝活性相關,因血凝素(纖突)具有型特異性,常用血凝抑制試驗(HI)對臨床分離株進行分型。
分類及自上個世紀50年代發現並成功分離腺病毒以來,已陸續發現了100餘個血清型,其中人腺病毒有52種,分為A、B、C、D、E和F六個亞群(subgroup)。基因治療常用的人的2型及5型腺病毒在血清學分類上均屬C亞群,在DNA序列上有95%的同源性。二者的增殖能力非常強,滴度通常可以達到109pfu (plaque forming unit)/ml,其在單個細胞中的基因組拷貝數可達104(約佔細胞總DNA的10%)。病毒顆粒比較穩定,通過CsCl梯度離心可以達到1010~1011pfu/ml,滿足動物實驗的要求。
腺病毒對呼吸道、胃腸道、尿道和膀胱、眼、肝臟等均可感染,人腺病毒約1/3的已知血清型通常與人類疾病相關,但一種血清型可引起不同的臨床疾患;相反,不同血清型也可引起同一種疾患。
腺病毒
1.急性發熱性咽喉炎 通常為嬰兒和兒童發病,由C組病毒引起,出現咳嗽,鼻塞、發熱和咽喉部潰瘍等癥狀,這些表現難以與其他病毒引起的輕型呼吸道感染鑒別。
2.咽結膜熱(pharyngoconjunctival fever)癥狀與急性發熱性咽喉炎相似,但常同時發生結膜炎。咽結膜熱有暴發流行傾向,如游泳池結膜炎,多由B組腺病毒3和7型所致,愈后尚好,一般無後遺症。
3.急性呼吸道疾病(acute respiratory diseases,ARD)這一綜合征由咽炎、發熱、咳嗽和全身不適為特點,常在軍隊的新兵中流行,多因突然緊張、勞累、聚集等所致。此感染多由腺病毒4、7型引起,也可見於3型。
4、肺炎 腺病毒肺炎約佔兒童期肺炎的10%,多由腺病毒3、7型引起;在青年人腺病毒肺炎的病死率為8%~10%;肺炎也是新兵急性呼吸道疾病的一種嚴重表現。
腺病毒致輕型眼部感染是呼吸道感染和咽喉炎的併發症。濾泡性結膜炎可由許多型腺病毒引起,類似於衣原體性結膜炎,而且為自限性。由腺病毒8、9和37型引起的角結膜炎為重型感染,具高傳染性,以急性結膜炎開始,擴至耳前淋巴結,隨後發生角膜炎。
腺病毒11、12型能引起兒童急性出血性膀胱炎,尿中出現病毒。37型可引起女性宮頸炎和男性尿道炎,常由性傳播感染。在免疫功能低下者可引起偶發或嚴重的病毒感染,尤其在器官移植病人中發生嚴重呼吸道感染和病毒性肝炎,多由1、5和7型腺病毒引起。艾滋病患者可感染多種血清型腺病毒,並能出現抗原性介於中間的雜合型毒株,而且常為致死性腺病毒感染。主要原因是腺病毒的E1A蛋白可反式激活HIV的轉錄,加速HIV的複製。臨床發現37%的艾滋病患者病毒性腹瀉是由腺病毒所致。
腺病毒感染后可獲得對同型的持久免疫力
與絕大多數呼吸道感染的病原相比,機體對腺病毒的再感染能產生有效的免疫。起保護作用的是體內產生的循環中和抗體。正常的健康成人一般也具有多型的抗體。約40%~60%的6~15歲的人具有1、2和5型中和抗體,但3、4和7型抗體很少。母親的抗體能保護嬰兒免除嚴重的腺病毒呼吸道感染。
腺病毒的生活周期可以分為兩個截然不同卻又不能割裂開來的兩個階段。第一階段包括腺病毒顆粒粘附和進入宿主細胞,將基因組釋放到宿主細胞核中,以及有選擇性地轉錄和翻譯早期基因。在這個階段,細胞為病毒基因組複製和腺病毒晚期基因表達並最終釋放成熟的感染顆粒,即第二階段,做好了準備。第一階段將在6~8個小時內完成,第二階段則更快,只需4~6個小時。
粘附和進入細胞
腺病毒感染細胞的過程是從腺病毒纖毛的頭節區粘附到細胞表面的特異性受體開始的。因為人腺病毒主要與柯薩奇B病毒共用一種受體,因此這種受體被稱為柯薩奇/腺病毒受體即CAR(coxsackie/adenovirus receptor)。接下來病毒纖毛基底部五鄰體表面的三肽RGD與細胞表面的αvβ3和αvβ5整合素結合,通過內吞作用將腺病毒內化到細胞中並進入溶酶體。在溶酶體的酸性環境下,腺病毒衣殼的構象將發生變化,被從溶酶體中釋放出來,躲過溶酶體的消化作用。最後,腺病毒顆粒轉位到細胞核,通過核孔將病毒DNA釋放到細胞核內。相對於脂質體轉染,腺病毒基因組進入細胞核是一個非常高效的過程,一般可以達到40%,前者雖然進入胞質的效率與後者相當,而DNA進入細胞核的效率卻只有後者的1/1000。
轉錄與複製
一旦病毒基因組進入細胞核,就將進行一系列的複雜而有序的逐級放大的剪切和轉錄過程。一般的,以病毒DNA開始複製為分界線,按轉錄時間的先後,將腺病毒基因大致區分為早期(E1~4)和晚期轉錄單位(L1~5)。各種腺病毒基因又可以進一步地分為更小的轉錄單位,如E1區可以進一步分為E1A和E1B,每個轉錄單位都至少有一個獨特的啟動子。腺病毒基因組進入細胞核后,細胞轉錄因子首先與E1A區上游的增強子結合,表達E1A蛋白,該蛋白的作用是調節細胞代謝,使病毒DNA更易於在細胞中複製。E1A蛋白還可以激活其他早期基因(E1B、E2A、E2B、E3和E4)的啟動子,其中E2B驅動另外三個與病毒複製有關的早期基因轉錄單位末端蛋白前體(pTP, precursor terminal protein)、單鏈DNA結合蛋白(ssDBP, single-stranded DNA binding proteins)以及DNA聚合酶(DNA pol, DNA polymerase)的表達,這三個基因的表達產物緊密地結合成一個複合物,與至少三種細胞蛋白相互作用,啟動病毒基因組的複製。
腺病毒
病毒基因組複製通常在感染后數小時開始,同時早期基因的轉錄和翻譯被關閉,晚期基因開始表達。大部分的晚期基因的轉錄是以一個共同的主要晚期啟動子(MLP, Major Late Promoter)調控的。實際上,MLP的活性與病毒基因組複製密切相關,有研究表明一旦腺病毒基因組開始複製MLP的活性將明顯增強,對此我們將另外行文詳述。晚期基因主要編碼腺病毒的結構蛋白。病毒結構蛋白在細胞核內聚集形成病毒衣殼,病毒的基因組被包裝進去,形成有感染能力的病毒顆粒,並最終裂解宿主細胞被釋放出去,完成腺病毒的生活周期。腺病毒有明顯的種屬特異性,人的野生型5型腺病毒(wtAd5)感染其他的非人類細胞(如鼠類細胞)后可以表達早期基因,基因組也可有一定程度的複製並能夠形成一些不成熟的病毒顆粒,卻不能形成成熟的病毒顆粒,也不能二次感染其他細胞。
對難於培養的腸道腺病毒,從糞便標本中粗提后,經電子顯微鏡或免疫電鏡觀察。
病毒分離與鑒定
1.分離培養 標本應儘早從感染部位採集。採集患者咽喉、眼分泌物,糞便和尿液等,加抗生素處理過夜,離心取上清接種敏感細胞(293、Hep-2或HeLa細胞等),37℃孵育后可觀察到典型CPE,即細胞變圓、團聚、有拉絲現象,最突出的表現是許多病變細胞聚在一起呈葡萄串狀。
2.病毒鑒定 用熒游標記的抗六鄰體抗體與分離培養細胞作用來鑒定腺病毒,也可用血凝抑制(hemoagglutination inhibition,HI)試驗或中和試驗(neutralization test,NT)檢測屬和組特異性抗原並鑒定病毒的血清型。
腺病毒可用Shell vial技術進行快速鑒定。病毒標本經抗生素和離心處理,取上清接種於有細胞的Shell vial培養瓶,孵育1~2天,用特異性六鄰體單克隆抗體對其抗原表位進行檢測。也可用病人鼻粘膜上皮脫落細胞直接染色檢測病毒抗原。
用DNA雜交或內切酶酶切等鑒定分離培養的病毒DNA;PCR可用於腺病毒感染的診斷,引物設計主要根據腺病毒六鄰體、VAI和VAII編碼區序列,能檢測所有血清型,而且其敏感性很高,能檢測某些病人潛在的腺病毒。用腺病毒41型BgIII-D片段作探針診斷腺病毒腹瀉,其檢出率可達80%。
常用血清學方法包括IF、CF、EIA、HI及NT等試驗,採取患者急性期和恢復期雙份血清進行檢測,若恢復期血清抗體效價比急性期增長4倍或以上,即有診斷意義。快速檢測血清可用ELISA法或乳膠凝集試驗。
常用來直接檢測腺病毒在呼吸道和胃腸道的感染,較快速且靈敏度較高。免疫熒光(尤其對呼吸道標本、咽拭子和活組織標本)和酶免疫分析(尤其對於糞便標本)是常用的方法,與細胞培養相比,免疫熒光所測腺病毒的靈敏性能提高40%~60%,其它直接測定抗原的方法包括免疫層析法和乳膠凝集法。研究證實,與細胞培養檢測方法相比,使免疫層析試劑盒所測定的靈敏度可達90%。
腺病毒的甲醛滅活疫苗已被用於某些人群的預防,而且將來有被用人二倍體細胞培養的減毒活疫苗所替代的可能。但因腺病毒對動物具有致癌作用,人們對全病毒疫苗的作用與安全性存有疑慮。此外加強游泳池和浴池水的消毒,可使水傳播性結膜炎爆發的危險性降至最小,在作眼的檢查時應嚴格無菌操作,對所用設備充分滅菌,也可控制流行性結膜炎的發生。對腺病毒感染的治療仍無有效藥物。
腺病毒基因產物的主要功能
E1區基因表達產物
可以進一步分為E1A和E1B。E1A主要由兩種成分構成,分別為289R(或13S)和243R(或12S)。這些E1A蛋白的主要功能是調節細胞代謝,使細胞對病毒複製更易感E1B19K與細胞Bcl-2基因的表達產物同源,可以通過滅活和清除Bax家族成員來防止細胞發生凋亡或壞死。E1B55K基因產物可以下調p53基因的轉錄水平,當然這種調節功能不是絕對的,其他一些腺病毒基因(如E4 orf6)也參與了這一過程。另外,E1B55K基因產物還與病毒複製、病毒晚期mRNA的轉錄以及病毒RNA的轉運有關。
E2區基因表達產物
可分為E2A和E2B。其中,E2A即DNA結合蛋白(DBP,DNA Binding Protein);E2B主要產物有兩種,分別是末端蛋白前體(pTP)和病毒DNA聚合酶(pol)。三種蛋白與至少三種細胞內的因子相互作用,啟動腺病毒DNA複製以及病毒晚期基因的轉錄和翻譯過程。
E3區基因表達產物
主要功能是破壞宿主的免疫防禦機制,而與病毒基因組的複製無關。E3基因的產物之一11.6Kd,由於可以在病毒感染的晚期裂解細胞並釋放病毒顆粒而被稱為腺病毒死亡蛋白(ADP,adenovirus death protein)。gp 19K蛋白可以在內質網上與MHC I類分子的重鏈結合阻止其轉運到細胞表面,並且可以延緩MHC I的表達。RID α&β以及14.7Kd可以抑制由TNF誘發的細胞凋亡,促進Fas降解,下調TNF受體水平。
E4區基因表達產物
E4區的基因產物通常被稱為orf 1~6/7,主要與病毒信使RNA的代謝有關。還有促進病毒DNA複製以及關閉宿主蛋白合成的功能。研究發現,一些E4產物可以與DNA激活的蛋白激酶結合,防止病毒DNA發生串聯。由於該激酶可以激活p53基因,因此可以認為一些E4區基因產物可以抑制細胞凋亡。許多E1B和E4基因產物都與拮抗E1A蛋白功能有關。比如,E4 orf4抑制E1A對E2F啟動子的激活;E4 orf3VA RNA使一些由腺病毒轉錄的非翻譯RNA,與腺病毒抵抗宿主細胞免疫有關。
宿主範圍廣對人致病性低
腺病毒載體系統可廣泛用於人類及非人類蛋白的表達。腺病毒可感染一系列哺乳動物細胞,因此在大多哺乳動物細胞和組織中均可用來表達重組蛋白。特別需要指出的是:腺病毒具有嗜上皮細胞性,而人類的大多數的腫瘤就是上皮細胞來源的。另外,腺病毒的複製基因和致病基因均已相當清楚,在人群中早已流行(70-80%成人體內都有腺病毒的中和抗體存在)。人類感染野生型腺病毒后僅產生輕微的自限性癥狀,且病毒唑治療有效。
在增殖和非增殖細胞中感染和表達基因
逆轉錄病毒只能感染增殖性細胞,因此DNA轉染不能在非增殖細胞中進行,而必須使細胞處於持續培養狀態。腺病毒則能感染幾乎所有的細胞類型,除了一些抗腺病毒感染的淋巴瘤細胞。腺病毒是研究原代非增殖細胞基因表達的最佳系統,它可以使轉化細胞和原代細胞中得到的結果直接進行對比。
能有效進行增殖 滴度高
腺病毒系統可產生10 到10 VP/ml,濃縮后可達10 VP/ml,這一特點使它非常適用於基因治療。
與人類基因同源
腺病毒載體系統一般應用人類病毒作為載體,以人類細胞作為宿主,因此為人類蛋白進行準確的翻譯后加工和適當的摺疊提供了一個理想的環境。大多數人類蛋白都可達到高水平表達並且具有完全的功能。
不整合到染色體中 無插入致突變性
逆轉錄病毒可隨機整合到宿主染色體,導致基因失活或激活癌基因。而腺病毒則除了卵細胞以外幾乎在所有已知細胞中都不整合到染色體中,因此不會幹擾其它的宿主基因。在卵細胞中整合單拷貝病毒則是產生具有特定特徵的轉基因動物的一個較好的系統。
能在懸浮培養液中擴增
293細胞可以適應懸浮培養,這一調整可使病毒大量擴增。大量事實證明懸浮293細胞可在1~20L的生物反應器中表達重組蛋白。
能同時表達多個基因
這是第一個可以在同一細胞株或組織中用來設計表達多個基因的表達系統。最簡單的方法是將含有兩個基因的雙表達盒插入腺病毒轉移載體中,或者用不同的重組病毒共轉染目的細胞株來分別表達一個蛋白。測定不同重組病毒的MOI比值可正確估計各重組蛋白的相對共表達情況。
正是由於具有以上一些優點,腺病毒被極其廣泛地應用於體外基因轉導、體內接種疫苗、和基因治療等各個領域。
腺病毒感染主要在冬春季流行,容易在幼兒園、學校和軍營新兵中暴發流行。一般來說,腺病毒主要通過呼吸道飛沫、眼分泌物,經呼吸道或接觸傳播;腸道感染主要通過消化道傳播。其預防措施和其他呼吸道、消化道傳染病預防相似,主要是勤洗手,勤消毒,避免接觸患者及其呼吸道飛沫。平常多飲水,多吃蔬菜和水果,注意鍛煉身體;室內多通風,保持室內環境清潔;冬春流行季節盡量少去人員密集的公共場所,外出時戴口罩,避免接觸病人,以防感染。
一旦發生急性發熱、咽喉疼痛和結膜炎的癥狀,要及早到醫院看病,早隔離、早治療。出現5人以上集體發病的情況要及時向所在地區防疫部門報告,及時採取有效的防控措施,避免疾病蔓延。在腺病毒流行季節,托幼機構上呼吸道感染患兒應回家隔離休息,以免造成傳播流行。患病後盡量在附近醫院就診,避免到病人較集中的大醫院觀察室輸液,以防造成交叉感染。出現嚴重咳嗽和呼吸困難癥狀多屬嚴重病例,應及時到醫院住院治療,以免延誤病情。
基本信息
- 中文名
- 腺病毒
- 拉丁學名
- adenovirus
- 屬性
- 沒有包膜的直徑為70~90nm
- 組成
- 252個殼粒呈廿面體排列構成
- 結構
- 基因組長約25-45kb
- 種類個數
- 已知有52種
- 含量
- 含13%DNA和87%的蛋白質