共找到5條詞條名為PCR的結果 展開
- 生物學的聚合酶鏈反應
- 主成分回歸分析
- 電子信息學的光電導繼電器
- 電子學的節目時鐘參考
- 計算機學的程序控制暫存器
PCR
生物學的聚合酶鏈反應
聚合酶鏈式反應(PCR)是一種用於放大擴增特定的DNA片段的分子生物學技術,它可看作是生物體外的特殊DNA複製,PCR的最大特點是能將微量的DNA大幅增加。因此,無論是化石中的古生物、歷史人物的殘骸,還是幾十年前兇殺案中兇手所遺留的毛髮、皮膚或血液,只要能分離出一丁點的DNA,就能用PCR加以放大,進行比對。這也是“微量證據”的威力之所在。由1983年美國Mullis首先提出設想,1985年由其發明了聚合酶鏈反應,即簡易DNA擴增法,意味著PCR技術的真正誕生。到如今2013年,PCR已發展到第三代技術。1976年,台灣科學家錢嘉韻,發現了穩定的Taq DNA聚合酶,為PCR技術發展也做出了基礎性貢獻。
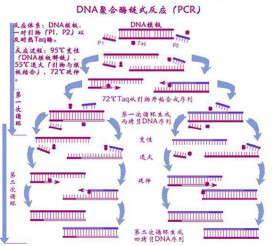
PCR是利用DNA在體外攝氏95°高溫時變性會變成單鏈,低溫(經常是60°C左右)時引物與單鏈按鹼基互補配對的原則結合,再調溫度至DNA聚合酶最適反應溫度(72°C左右),DNA聚合酶沿著磷酸到五碳糖(5'-3')的方向合成互補鏈。基於聚合酶製造的PCR儀實際就是一個溫控設備,能在變性溫度,復性溫度,延伸溫度之間很好地進行控制。
Khorana (1971)等最早提出核酸體外擴增的設想:“經DNA變性,與合適的引物雜交,用DNA聚合酶延伸引物,並不斷重複該過程便可合成tRNA基因。”
但由於當時基因序列分析方法尚未成熟,熱穩定DNA聚合酶尚未報道以及引物合成的困難,這種想法似乎沒有實際意義。加上分子克隆技術的出現提供了一種克隆和擴增基因的途徑,所以Khorana的設想被人們遺忘了。
1985年,Kary Mullis在Cetus公司工作期間,發明了PCR。Mullis要合成DNA引物來進行測序工作,卻常為沒有足夠多的模板DNA而煩惱。
1983年4月的一個星期五晚上,他開車去鄉下別墅的路上,猛然閃現出“多聚酶鏈式反應”的想法。
1983年12月,Mullis用同位素標記法看到了10個循環后的49 bp長度的第一個PCR片段;
1985年10月25日申請了PCR的專利,1987年7月28日批准(專利號4,683,202 ),Mullis是第一發明人;
1985年12月20日在Science雜誌上發表了第一篇PCR的學術論文,Mullis是共同作者;
1986年5月,Mullis在冷泉港實驗室做專題報告,全世界從此開始學習PCR的方法。
DNA的半保留複製是生物進化和傳代的重要途徑。雙鏈DNA在多種酶的作用下可以變性解旋成單鏈,在DNA聚合酶的參與下,根據鹼基互補配對原則複製成同樣的兩分子拷貝。在實驗中發現,DNA在高溫時也可以發生變性解鏈,當溫度降低后又可以復性成為雙鏈。因此,通過溫度變化控制DNA的變性和復性,加入設計引物,DNA聚合酶、dNTP就可以完成特定基因的體外複製。
但是,DNA聚合酶在高溫時會失活,因此,每次循環都得加入新的DNA聚合酶,不僅操作煩瑣,而且價格昂貴,制約了PCR技術的應用和發展。
耐熱DNA聚合酶-Taq酶的發現對於PCR的應用有里程碑的意義,該酶可以耐受90℃以上的高溫而不失活,不需要每個循環加酶,使PCR技術變得非常簡捷、同時也大大降低了成本,PCR技術得以大量應用,並逐步應用於臨床。
PCR技術的基本原理類似於DNA的天然複製過程,其特異性依賴於與靶序列兩端互補的寡核苷酸引物。PCR由變性-退火-延伸三個基本反應步驟構成:①模板DNA的變性:模板DNA經加熱至93℃左右一定時間后,使模板DNA雙鏈或經PCR擴增形成的雙鏈DNA解離,使之成為單鏈,以便它與引物結合,為下輪反應作準備;②模板DNA與引物的退火(復性):模板DNA經加熱變性成單鏈后,溫度降至55℃左右,引物與模板DNA單鏈的互補序列配對結合;③引物的延伸:DNA模板-引物結合物在72℃、DNA聚合酶(如TaqDNA聚合酶)的作用下,以dNTP為反應原料,靶序列為模板,按鹼基互補配對與半保留複製原理,合成一條新的與模板DNA鏈互補的半保留複製鏈,重複循環變性-退火-延伸三過程就可獲得更多的“半保留複製鏈”,而且這種新鏈又可成為下次循環的模板。每完成一個循環需2~4分鐘,2~3小時就能將待擴目的基因擴增放大幾百萬倍。
| 10×擴增緩衝液 | 10μl |
| 4種dNTP混合物(終濃度) | 各100~250μmol/L |
| 引物(終濃度) | 各5~20μmol/L |
| 模板DNA | 0.1~2μg |
| Taq DNA聚合酶 | 5~10 U |
| Mg(終濃度) | 1~3mmol/L |
| 補加雙蒸水 | 100 μl |
其中dNTP、引物、模板DNA、Taq DNA聚合酶以及Mg 的加量(或濃度)可根據實驗調整,以上表格只提供大致參考值。
PCR反應五要素:參加PCR反應的物質主要有五種即:引物(PCR引物為DNA片段,細胞內DNA複製的引物為一段RNA鏈)、酶、dNTP、模板和緩衝液(其中需要Mg )。
引物有多種設計方法,由PCR在實驗中的目的決定,但基本原則相同。PCR所用的酶主要有兩種來源:Taq和Pfu,分別來自兩種不同的噬熱菌。其中Taq擴增效率高但易發生錯配。Pfu擴增效率弱但有糾錯功能。所以實際使用時根據需要必須做不同的選擇。
模板即擴增用的DNA,可以是任何來源,但有兩個原則,第一純度必須較高,第二濃度不能太高以免抑制。
緩衝液的成分最為複雜,除水外一般包括四個有效成分:緩衝體系,一般使用HEPES或MOPS緩衝體系;一價陽離子,一般採用鉀離子,但在特殊情況下也可使用銨根離子;二價陽離子,即鎂離子,根據反應體系確定,除特殊情況外不需調整;輔助成分,常見的有DMSO、甘油等,主要用來保持酶的活性和幫助DNA解除纏繞結構。
PCR反應中有兩條引物,即5′端引物和3′引物。設計引物時以一條DNA單鏈為基準(常以信息鏈為基準),5′端引物與位於待擴增片段5′端上的一小段DNA序列相同;3′端引物與位於待擴增片段3′端的一小段DNA序列互補。
(1)引物設計的基本原則
• 引物長度:15-30bp,常用為20bp左右。
• 引物鹼基:G+C含量以40-60%為宜,G+C太少擴增效果不佳,G+C 過多易出現非特異條帶。ATGC最好隨機分佈,避免5個以上的嘌呤或嘧啶核苷酸的成串排列參照。
• 引物內部不應出現互補序列。
• 兩個引物之間不應存在互補序列,尤其是避免3 ′端的互補重疊。
• 引物與非特異擴增區的序列的同源性不要超過70%,引物3′末端連續8個鹼基在待擴增區以外不能有完全互補序列,否則易導致非特異性擴增。
• 引物3‘端的鹼基,特別是最末及倒數第二個鹼基,應嚴格要求配對,最佳選擇是G和C。
• 引物的5′端可以修飾。如附加限制酶位點,引入突變位點,用生物素、熒光物質、地高辛標記,加入其它短序列,包括起始密碼子、終止密碼子等。
(2)引物設計軟體
• Primer Premier5.0 (自動搜索)
• vOligo6 (引物評價)
• vVector NTI Suit
• vDNAsis
• vOmiga
• vDNAstar
• vPrimer3 (在線服務)
PCR的模板可以是DNA,也可以是RNA。
模板的取材主要依據PCR的擴增對象,可以是病原體標本如病毒、細菌、真菌等。也可以是病理生理標本如細胞、血液、羊水細胞等。法醫學標本有血斑、精斑、毛髮等。
標本處理的基本要求是除去雜質,並部分純化標本中的核酸。多數樣品需要經過SDS和蛋白酶K處理。難以破碎的細菌,可用溶菌酶加EDTA處理。所得到的粗製DNA,經酚、氯仿抽提純化,再用乙醇沉澱後用作PCR反應模板。
• PCR反應的緩衝液 提供合適的酸鹼度與某些離子
• 鎂離子濃度 總量應比dNTPs的濃度高,常用1.5mmol/L
• 底物濃度 dNTP以等摩爾濃度配製,20~200umol/L
• TaqDNA聚合酶 2.5U(100ul)
• 引物 濃度一般為0.1 ~ 0.5umol/L
• 反應溫度和循環次數
變性溫度和時間 95℃,30s
退火溫度和時間 低於引物Tm值5 ℃左右,一般在45~55℃
延伸溫度和時間 72℃,1min/kb(10kb內)
Tm值=4(G+C) +2(A+T)
循環次數:一般為25 ~ 30次。循環數決定PCR擴增的產量。模板初始濃度低,可增加循環數以便達到有效的 擴增量。但循環數並不是可以無限增加的。一般循環數為30個左右,循環數超過30個以後,DNA聚合酶活性逐漸達到飽和,產物的量不再隨循環數的增加而增加,出現了所謂的“平台期”。
模板DNA完全變性與PCR酶的完全激活對PCR能否成功至關重要,建議加熱時間參考試劑說明書,一般未修飾的Taq酶激活時間為兩分鐘。
循環中一般95℃,30秒足以使各種靶DNA序列完全變性,可能的情況下可縮短該步驟時間。變性時間過長損害酶活性,過短靶序列變性不徹底,易造成擴增失敗。
退火溫度需要從多方面去決定,一般根據引物的Tm值為參考,根據擴增的長度適當下調作為退火溫度。然後在此次實驗基礎上做出預估。退火溫度對PCR的特異性有較大影響。
引物延伸一般在72℃進行(Taq酶最適溫度)。但在擴增長度較短且退火溫度較高時,本步驟可省略延伸時間隨擴增片段長短而定,一般推薦在1000bp以上,含Pfu及其衍生物的衍生設定為1min/kbp。
大多數PCR含25-35循環,過多易產生非特異擴增。
在最後一個循環后,反應在72℃維持10-30分鐘.使引物延伸完全,並使單鏈產物退火成雙鏈。
標準的PCR過程分為三步:
DNA變性:(90℃-96℃):雙鏈DNA模板在熱作用下,氫鍵斷裂,形成單鏈DNA
退火:(60℃-65℃):系統溫度降低,引物與DNA模板結合,形成局部雙鏈。
延伸:(70℃-75℃):在Taq酶(在72℃左右,活性最佳)的作用下,以dNTP為原料,從引物的3′端開始以從5′→3′端的方向延伸,合成與模板互補的DNA鏈。
每一循環經過變性、退火和延伸,DNA含量即增加一倍。如圖所示:現在有些PCR因為擴增區很短,即使Taq酶活性不是最佳也能在很短的時間內複製完成,因此可以改為兩步法,即退火和延伸同時在60℃-65℃間進行,以減少一次升降溫過程,提高了反應速度。
PCR反應擴增出了高的拷貝數,下一步檢測就成了關鍵。熒光素(溴化乙錠,EB)染色凝膠電泳是最常用的檢測手段。電泳法檢測特異性是不太高的,因此引物兩聚體等非特異性的雜交體很容易引起誤判。但因為其簡捷易行,成為了主流檢測方法。近年來以熒光探針為代表的檢測方法,有逐漸取代電泳法的趨勢。
PCR反應的特異性決定因素為:
①引物與模板DNA特異正確的結合;
②鹼基配對原則;
③Taq DNA聚合酶合成反應的忠實性;
④靶基因的特異性與保守性。
其中引物與模板的正確結合是關鍵。引物與模板的結合及引物鏈的延伸是遵循鹼基配對原則的。聚合酶合成反應的忠實性及TaqDNA聚合酶耐高溫性,使反應中模板與引物的結合(復性)可以在較高的溫度下進行,結合的特異性大大增加,被擴增的靶基因片段也就能保持很高的正確度。再通過選擇特異性和保守性高的靶基因區,其特異性程度就更高。
PCR產物的生成量是以指數方式增加的,能將皮克(pg=10-12)量級的起始待測模板擴增到微克(μg=-6)水平。能從100萬個細胞中檢出一個靶細胞;在病毒的檢測中,PCR的靈敏度可達3個RFU(空斑形成單位);在細菌學中最小檢出率為3個細菌。
PCR反應用耐高溫的Taq DNA聚合酶,一次性地將反應液加好后,即在DNA擴增液和水浴鍋上進行變性-退火-延伸反應,一般在2~4 小時完成擴增反應。擴增產物一般用電泳分析,不一定要用同位素,無放射性污染、易推廣。
不需要分離病毒或細菌及培養細胞,DNA 粗製品及RNA均可作為擴增模板。可直接用臨床標本如血液、體腔液、洗嗽液、毛髮、細胞、活組織等DNA擴增檢測。
PCR產物的電泳檢測時間一般為48h以內,有些最好於當日電泳檢測,大於48h后帶型不規則甚至消失。
不出現擴增條帶。PCR反應的關鍵環節有①模板核酸的製備,②引物的質量與特異性,③酶的質量及溴乙錠的使用, ④PCR循環條件。尋找原因亦應針對上述環節進行分析研究。
模板:①模板中含有雜蛋白質,②模板中含有Taq酶抑製劑,③模板中蛋白質沒有消化除凈,特別是染色體中的組蛋白,④在提取製備模板時丟失過多,或吸入酚。⑤模板核酸變性不徹底。在酶和引物質量好時,不出現擴增帶,極有可能是標本的消化處理,模板核酸提取過程出了毛病,因而要配製有效而穩定的消化處理液,其程序亦應 固定不宜隨意更改。
需注意的是有時忘加Taq酶或溴乙錠。引物:引物質量、引物的濃度、兩條引物的濃度是否對稱,是PCR失敗或擴增條帶不理想、容易彌散的常見原因。有些批號的引物合成質量有問題,兩條引物一條濃度高,一條濃度低,造成低效率的不對稱擴增,對策為:①選定一個好的引物合成單位。②引物的濃度不僅要看OD值,更要注重引物原液做瓊脂糖凝膠電泳,一定要有引物條帶出現,而且兩引物帶的亮度應大體一致,如一條引物有條帶,一條引物無條帶,此時做PCR有可能失敗,應和引物合成單位協商解決。如一條引物亮度高,一條亮度低,在稀釋引物時要平衡其濃度。③引物應高濃度小量分裝保存,防止多次凍融或長期放冰箱冷藏部分,導致引物變質
降解失效。④引物設計不合理,如引物長度不夠,引物之間形成二聚體等。
Mg 濃度:Mg 離子濃度對PCR擴增效率影響很大,濃度過高可降低PCR擴增的特 異性,濃度過低則影響PCR擴增產量甚至使PCR擴增失敗而不出擴增條帶。
反應體積的改變:通常進行PCR擴增採用的體積為20ul、30ul、50ul。或100ul,應用多 大體積進行PCR擴增,是根據科研和臨床檢測不同目的而設定,在做小體積如20ul 后,再做大體積時,一定要模索條件,否則容易失敗。
物理原因:變性對PCR擴增來說相當重要,如變性溫度低,變性時間短,極有可能出現假陰性;退火溫度過低,可致非特異性擴增而降低特異性擴增效率退火溫度過高影響引物與模板的結合而降低PCR擴增效率。有時還有必要用標準的溫度計,檢測一下擴增儀或水溶鍋內的變性、退火和延伸溫度,這也是PCR失敗的原因之一。
靶序列變異:如靶序列發生突變或缺失,影響引物與模板特異性結合,或因靶序列某 段缺失使引物與模板失去互補序列,其PCR擴增是不會成功的。
出現的PCR擴增條帶與目的靶序列條帶一致,有時其條帶更整齊,亮度更高。
引物設計不合適:選擇的擴增序列與非目的擴增序列有同源性,因而在進行PCR擴增時,擴增出的PCR產物為非目的性的序列。靶序列太短或引物太短,容易出現假陽性。需重新設計引物。
靶序列或擴增產物的交叉污染:這種污染有兩種原因:一是整個基因組或大片段的交叉污染,導致假陽性。這種假陽性可用以下方法解決:操作時應小心輕柔,防止將靶序列吸入加樣槍內或濺出離心管外。除酶及不能耐高溫的物質外,所有試劑或器材均應高壓消毒。所用離心管及樣進槍頭等均應一次性使用。必要時,在加標本前,反應管和試劑用紫外線照射,以破壞存在的核酸。二是空氣中的小片段核酸污染,這些小片段比靶序列短,但有一定的同源性。可互相拼接,與引物互補后,可擴增出PCR產物,而導致假陽性的產生,可用巢式PCR方法來減輕或消除。
出現非特異性擴增帶:
PCR擴增后出現的條帶與預計的大小不一致,或大或小,或者同時出現特異性擴增帶 與非特異性擴增帶。非特異性條帶的出現,其原因:一是引物與靶序列不完全互補、或引物聚合形成二聚體。二是Mg 離子濃度過高、退火溫度過低,及PCR循環次數過多有關。其次是酶的質和量,往往一些來源的酶易出現非特異條帶而另一來源的酶則不出現,酶量過多有時也會出現非特異性擴增。其對策有:必要時重新設計引 物。減低酶量或調換另一來源的酶。降低引物量,適當增加模板量,減少循環次數。適當提高退火溫度或採用二溫度點法(93℃變性,65℃左右退火與延伸)。
出現片狀拖帶或塗抹帶:
PCR擴增有時出現塗抹帶或片狀帶或地毯樣帶。其原因往往由於酶量過多或酶的質量差,dNTP濃度過高,Mg 濃度過高,退火溫度過低,循環次數過多引起。其對策有:減少酶量,或調換另一來源的酶。②減少dNTP的濃度。適當降低Mg 濃 度。增加模板量,減少循環次數。
用於治療感染性疾病,腫瘤及遺傳病。
PCR在醫學檢驗學中最有價值的應用領域就是對感染性疾病的診斷。理論上,只要樣本有一個病原體存在,PCR就可以檢測到。一般實驗室也能檢出10~100基因拷貝,而目前病原體抗原檢測方法一般需要105-7個病原體才可檢測到。PCR對病原體的檢測解決了免疫學檢測的“窗口期”問題,可判斷疾病是否處於隱性或亞臨床狀態。
定量PCR研究資料已表明,病原體數量與感染性疾病病情的輕重程度、傳染性及治療效果均有相關性。許多研究表明,人類免疫缺陷病毒(HIV)感染后,潛伏期長短和臨床癥狀輕重與血液中的病毒量顯著相關;也有研究表明,HIV病毒載量低於一定值時,沒有傳染性。
在乙型肝炎病毒、丙型肝炎病毒定量研究中發現,病毒的數量與某些藥物的療效相關。例如,干擾素治療對肝炎病毒高拷貝者不敏感,低拷貝者敏感;而有些藥物則具有顯著降低病毒高拷貝的作用。
癌基因的表達增加和突變,在許多腫瘤早期和良性的階段就可出現。PCR技術不但能有效的檢測基因的突變,而且能準確檢測癌基因的表達量,可據此進行腫瘤早期診斷、分型、分期和預后判斷。
幾乎所有慢性骨髓性白血病患者都可檢測到原癌基因易位導致的BCR/ABL融合基因形成,定量PCR技術可通過檢測BCR/ABL融合基因的表達確定微量殘餘惡性細胞存在的數量,以此作為治療效果和估計複發的危險性的依據。
PCR反應的最大特點是具有較大擴增能力與極高的靈敏性,但令人頭痛的問題是易污染,極其微量的污染即可造成假陽性的產生。
1、標本間交叉污染:標本污染主要有收集標本的容器被污染,或標本放置時,由於密封不嚴溢於容器外,或容器外粘有標本而造成相互間交叉污染;標本核酸模板在提取過程中,由於吸樣槍污染導致標本間污染;有些微生物標本尤其是病毒可隨氣溶膠或形成氣溶膠而擴散,導致彼此間的污染。
2、PCR試劑的污染:主要是由於在PCR試劑配製過程中,由於加樣槍、容器、雙蒸水及其它溶液被PCR核酸模板污染。
3、PCR擴增產物污染:這是PCR反應中最主要最常見的污染問題。因為PCR產物拷貝量大(一般為1013拷貝/ml),遠遠高於PCR檢測數個拷貝的極限,所以極微量的PCR產物污染,就可形成假陽性。還有一種容易忽視,最可能造成PCR產物污染的形式是氣溶膠污染。在空氣與液體面摩擦時就可形成氣溶膠,在操作時比較劇烈地搖動反應管,開蓋時、吸樣時及污染進樣槍的反覆吸樣都可形成氣溶膠而污染。據計算一個氣溶膠顆粒可含48000拷貝,因而由其造成的污染是一個值得特別重視的問題。
4、實驗室中克隆質粒的污染:在分子生物學實驗室及某些用克隆質粒做陽性對照的檢驗室,這個問題也比較常見。因為克隆質粒在單位容積內含量相當高,另外在純化過程中需用較多的用具及試劑,而且在活細胞內的質粒,由於活細胞的生長繁殖的簡便性及具有很強的生命力。其污染可能性也很大。
一個好的實驗室,要時刻注意污染的監測,考慮有無污染是什麼原因造成的污染,以便採取措施,防止和消除污染。
1、陽性對照:在建立PCR反應實驗室及一般的檢驗單位都應設有PCR陽性對照,它是PCR反應是否成功、產物條帶位置及大小是否合乎理論要求的一個重要的參考標誌。陽性對照要選擇擴增度中等、重複性好,經各種鑒定是該產物的標本,如以重組質粒為陽性對照,其含量宜低不宜高(100個拷貝以下)。但陽性對照尤其是重組質粒及高濃度陽性標本,其對檢測或擴增樣品污染的可能性很大。因而當某一PCR試劑經自己使用穩定,檢驗人員心中有數時,在以後的實驗中可免設陽性對照。
2、陰性對照:每次PCR實驗務必做陰性對照。它包括:
(1)標本對照:被檢的標本是血清就用鑒定后的正常血清作對照;被檢的標本是組織細胞就用相應的組織細胞作對照。
(2)試劑對照:在PCR試劑中不加模板DNA或RNA,進行PCR擴增,以監測試劑是否污染。
3、重複性試驗。
4、選擇不同區域的引物進行PCR擴增。
基本信息
- 中文名
- 生物學的聚合酶鏈式反應
- 外文名
- Polymerase Chain Reaction
- 別名
- PCR
- 提出者
- Mullis
- 屬性
- 鏈式反應
- 目前狀態
- 第三代技術
- 提出人
- 穆利斯