共找到2條詞條名為基因突變的結果 展開
- 基因組DNA分子發生的突然的、可遺傳的變異現象
- 生物學名詞
基因突變
基因組DNA分子發生的突然的、可遺傳的變異現象
基因組DNA分子發生的突然的、可遺傳的變異現象(gene mutation)。從分子水平上看,基因突變是指基因在結構上發生鹼基對組成或排列順序的改變。基因雖然十分穩定,能在細胞分裂時精確地複製自己,但這種穩定性是相對的。在一定的條件下基因也可以從原來的存在形式突然改變成另一種新的存在形式,就是在一個位點上,突然出現了一個新基因,代替了原有基因,這個基因叫做突變基因。於是後代的表現中也就突然地出現祖先從未有的新性狀。
基因突變首先由T.H.摩爾根於1910年在果蠅中發現。H.J.馬勒於1927年、L.J.斯塔德勒於1928年分別用X射線等在果蠅、玉米中最先誘發了突變。1947年C.奧爾巴克首次使用了化學誘變劑,用氮芥誘發了果蠅的突變。1943年S.E.盧里亞和M.德爾布呂克最早在大腸桿菌中證明對噬菌體抗性的出現是基因突變的結果。接著在細菌對於鏈黴素和磺胺葯的抗性方面獲得同樣的結論。於是基因突變這一生物界的普遍現象逐漸被充分認識,基因突變的研究也進入了新的時期。1949年光復活作用發現后,DNA損傷修復的研究也迅速推進。這些研究結果說明基因突變並不是一個單純的化學變化,而是一個和一系列酶的作用有關的複雜過程。
1958年S.本澤發現噬菌體T4的rⅡ基因中有特別容易發生突變的位點──熱點,指出一個基因的某一對核苷酸的改變和它所處的位置有關。
1959年E.佛里茲提出基因突變的鹼基置換理論,1961年F.H.C.克里克等提出移碼突變理論(見遺傳密碼)。隨著分子遺傳學的發展和DNA核苷酸順序分析等技術的出現,已能確定基因突變所帶來的DNA分子結構改變的類型,包括某些熱點的分子結構,並已經能夠進行定向誘變。
鹼基置換示意圖
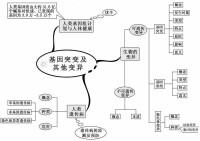
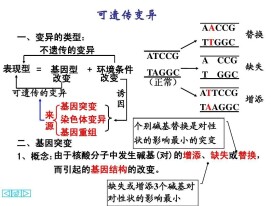
按照表型效應,突變型可以區分為形態突變型、生化突變型以及致死突變型等。這樣的區分並不涉及突變的本質,而且也不嚴格。因為形態的突變和致死的突變必然有它們的生物化學基礎,所以嚴格地講一切突變型都是生物化學突變型。根據鹼基變化的情況,基因突變一般可分為鹼基置換突變(base substitution和移碼突變(frameshift mutation)兩大類。
鹼基置換突變(subsititution)
BU誘發的突變
n)。嘌呤取代嘧啶或嘧啶取代嘌呤的突變則稱為顛換(transversion)。由於DNA分子中有四種鹼基,故可能出現4種轉換和8種顛換(見上圖)。在自然發生的突變中,轉換多於顛換。
亞硝胺誘發的突變
鹼基對的轉換也可由一些化學誘變劑誘變所致。例如,亞硝酸類能使胞嘧啶(C)氧化脫氨變成尿嘧啶(U),在下一 次複製中,U不與G配對,而與A配對;複製結果C-G變為T-A(見右圖)。又如,烷化劑中的芥子氣和硫酸二乙酯可使G發生乙基化,成為烷基化鳥嘌呤(mG),結果,mG不與C配對,而與T配對,經過複製,G-C變為A-T。
移碼突變(translocation)
指DNA片段中某一位點插入或丟失一個或幾個(非3或3的倍數)鹼基對時,造成插入或丟失位點以後的一系列編碼順序發生錯位的一種突變。它可引起該位點以後的遺傳信息都出現異常。發生了移碼突變的基因在表達時可使組成多肽鏈的氨基酸序列發生改變,從而嚴重影響蛋白質或酶的結構與功能。吖啶類誘變劑如原黃素、吖黃素、吖啶橙等由於分子比較扁平,能插入到DNA分子的相鄰鹼基對之間。如在DNA複製前插入,會造成1個鹼基對的插入;若在複製過程中插入,則會造成1個鹼基對的缺失,兩者的結果都引起移碼突變。
缺失突變(deletion)
基因也可以因為較長片段的DNA的缺失而發生突變。缺失的範圍如果包括兩個基因,那麼就好象兩個基因同時發生突變,因此又稱為多位點突變。由缺失造成的突變不會發生回復突變。所以嚴格地講,缺失應屬於染色體畸變。
插入突變(insertion)
一個基因的DNA中如果插入一段外來的DNA,那麼它的結構便被破壞而導致突變。大腸桿菌的噬菌體Mu-1和一些插入順序(IS)以及轉座子(見轉座因子)都是能夠轉移位置的遺傳因子,當它們轉移到某一基因中時,便使這一基因發生突變。許多轉座子上帶有抗藥性基因,當它們轉移到某一基因中時,一方面引起突變,另一方面使這一位置上出現一個抗藥性基因。插入的DNA分子可以通過切離而失去,準確的切離可以使突變基因回復成為野生型基因。這一事件的出現頻率並不由於誘變劑的處理而提高。
不論是真核生物還是原核生物的突變,也不論是什麼類型的突變,都具有隨機性、低頻性和可逆性等共同的特性。
普遍性
基因突變在自然界各物種中普遍存在。
隨機性
T.H.摩爾根在飼養的許多紅色複眼的果蠅中偶然發現了一隻白色複眼的果蠅。這一事實說明基因突變的發生在時間上、在發生這一突變的個體上、在發生突變的基因上,都是隨機的。以後在高等植物中所發現的無數突變都說明基因突變的隨機性。在細菌中則情況遠為複雜。在含有某一種藥物的培養基中培養細菌時往往可以得到對於這一藥物具有抗性的細菌,因此曾經認為細菌的抗藥性的產生是藥物引起的,是定向的適應而不是隨機的突變。S.盧里亞和M.德爾布呂克在1943年首先用波動測驗方法證明在大腸桿菌中的抗噬菌體細菌的出現和噬菌體的存在無關。J.萊德伯格等在1952年又用印影接種方法證實了這一論點。方法是把大量對於藥物敏感的細菌塗在不含藥物的培養基表面,把這上面生長起來的菌落用一塊滅菌的絲絨作為接種工具印影接種到含有某種藥物的培養基表面,使得兩個培養皿上的菌落的位置都一一對應。根據后一培養基表面生長的個別菌落的位置,可以在前一培養皿上找到相對應的菌落。在許多情況下可以看到這些菌落具有抗藥性。由於前一培養基是不含葯的,因此這一實驗結果非常直觀地說明抗藥性的出現不依賴於藥物的存在,而是隨機突變的結果,只不過是通過藥物將它們檢出而已。
稀有性
在第一個突變基因發現時,不是發現若干白色複眼果繩而是只發現一隻,說明突變是極為稀有的,也就是說野生型基因以極低的突變率發生突變(一些有代表性的基因突變率見表)。在有性生殖的生物中,突變率用每一配子發生突變的概率,也就是用一定數目配子中的突變型配子數表示。在無性生殖的細菌中,突變率用每一細胞世代中每一細菌發生突變的概率,也就是用一定數目的細菌在分裂一次過程中發生突變的次數表示。據估計,在高等生物中,大約10^5~10^8個生殖細胞中,才會有1個生殖細胞發生基因突變。雖然基因突變的頻率很低,但是當一個種群內有許多個體時,就有可能產生各種各樣的隨機突變,足以提供豐富的可遺傳的變異。
可逆性
野生型基因經過突變成為突變型基因的過程稱為正向突變。正向突變的稀有性說明野生型基因是一個比較穩定的結構。突變基因又可以通過突變而成為野生型基因,這一過程稱為回復突變。從表中同樣可以看到回復突變是難得發生的,說明突變基因也是一個比較穩定的結構。不過,正向突變率總是高於回復突變率,這是因為一個野生型基因內部的許多位置上的結構改變都可以導致基因突變,但是一個突變基因內部只有一個位置上的結構改變才能使它恢復原狀。
少利多害性
一般基因突變會產生不利的影響,被淘汰或是死亡,但有極少數會使物種增強適應性。
不定向性
例如控制黑毛A基因可能突變為控制白毛的a+或控制綠毛的a-基因。
有益性

解釋了一個鳥的基因突變或進化后的明顯區別
一般,基因突變後身體會發出抗體或其他修復體進行自行修復。可是有一些突變是不可迴轉性的。突變可能導致立即死亡,也可以導致慘重後果,如器官無法正常運作,DNA嚴重受損,身體免疫力低下等。如果是有益突變,可能會發生奇迹,如身體分泌中特殊變種細胞來保護器官,身體,或在一些沒有受骨骼保護的部位長出骨骼。基因與DNA就像是每個人的身份證,可他又是一個人的先知,因為它決定著身體的衰老、病變、死亡的時間。
獨立性
某一基因位點的一個等位基因發生突變,不影響另一個等位基因,即等位基因中的兩個基因不會同時發生突變。
①隱性突變:當代不表現,F2代表現。
②顯性突變:當代表現,與原性狀並存,形成鑲嵌現象或嵌合體。
重演性
同一生物不同個體之間可以多次發生同樣的突變。
基因突變
響蛋白質或酶的生物功能,使機體的表型出現異常。鹼基突變對多肽鏈中氨基酸序列的影響一般有下列幾種類型。
同義突變(same sense mutation):鹼基置換后,雖然每個密碼子變成了另一個密碼子,但由於密碼子的簡併性,因而改變前、后密碼子所編碼的氨基酸不變,故實際上不會發生突變效應。例如,DNA分子模板鏈中GCG的第三位G被A取代,變為GCA,則mRNA中相應的密碼子CGC就變為CGU,由於CGC和CGU都是編碼精氨酸的密碼子,故突變前後的基因產物(蛋白質)完全相同。同義突變約佔鹼基置換突變總數的25﹪。
錯義突變(missense mutation):鹼基對的置換使mRNA的某一個密碼子變成編碼另一種氨基酸的密碼子的突變稱為錯義突變。錯義突變可導致機體內某種蛋白質或酶在結構及功能發生異常,從而引起疾病。如人類正常血紅蛋白β鏈的第六位是谷氨酸,其密碼子為GAA或GAG,如果第二個鹼基A被U替代,就變成GUA或GUG,谷氨酸則被纈氨酸所替代,形成異常血紅蛋白HbS,導致個體產生鐮形細胞貧血,產生了突變效應。
基因突變
,產生沒有生物活性的多肽片段,稱為無義突變。例如,DNA分子中的ATG中的G被T取代時,相應mRNA鏈上的密碼子便從UAC變為UAA,因而使翻譯就此停止,造成肽鏈縮短。這種突變在多數情況下會影響蛋白質或酶的功能。
終止密碼突變(terminator codon mutation):基因中一個終止密碼突變為編碼某個氨基酸的密碼子的突變稱為終止密碼突變。由於肽鏈合成直到下一個終止密碼出現才停止,因而合成了過長的多肽鏈,故也稱為延長突變。例如,人血紅蛋白α鏈突變型Hb Constant Spring比正常人α珠蛋白鏈多了31個氨基酸。
外因
基因突變
化學因素:亞硝酸、黃麴黴素、鹼基類似物等。
生物因素:某些病毒和細菌等。
內因
DNA複製過程中,基因內部的脫氧核苷酸的數量、順序、種類發生了局部改變從而改變了遺傳信息
對於人類來講,基因突變可以是有用的也可以是有害的。

基因突變
變的有用的方面。在化學誘變劑發現以前,植物育種工作主要採用輻射作為誘變劑;化學誘變劑發現以後,誘變手段便大大地增加了。在微生物的誘變育種工作中,由於容易在短時間中處理大量的個體,所以一般只是要求誘變劑作用強,也就是說要求它能產生大量的突變。對於難以在短時間內處理大量個體的高等植物來講,則要求誘變劑的作用較強,效率較高並較為專一。所謂效率較高便是產生更多的基因突變和較少的染色體畸變。所謂專一便是產生特定類型的突變型。以色列培育“彩色青椒”關鍵技術就是把青椒種子送上太空,使其在完全失重狀態下發生基因突變來育種。

基因突變
的或不育的子代。
多數突變對於生物本身來講是有害的,人類的癌症的發生也和基因突變有密切的關係,因此環境中的誘變物質的檢測已成為公共衛生的一項重要任務。
從基因突變的性質來看,檢測方法分為顯性突變法、隱性突變法和回復突變法三類。①顯性致死突變法,用待測物質處理雄性小鼠,使處理的雄鼠和未處理的雌鼠交配,觀察母鼠子宮中的死胎數,死胎數愈多則說明誘發的顯性致死突變愈多。這一方法適用於慢性處理,其優點是可靠性較大,而且測試對象是哺乳動物。缺點是不能區別出藥物對遺傳物質的誘變作用和對於胚胎髮育的其他毒理效應。②隱性突變法,一般採用某些隱性突變基因呈雜合狀態的動植物作為測試對象,如果經某種藥物處理后出現這一隱性性狀,便說明這一藥物誘發了這一隱性突變。小鼠中有多個隱性突變基因呈雜合狀態的品系,可以用它來同時測定幾個座位上誘發的基因突變。這一方法的優點是所測得的是哺乳動物中的基因突變,缺點是靈敏度較低,而且必須具備特殊的動植物品系,實驗周期也較長。CIB法是用果蠅作為測試對象的一種檢測方法。主要用來檢測X染色體上發生的隱性致死突變。果蠅的生活周期較短,所以這一方法的實驗周期也較短。③回復突變法,一種根據回復突變誘發頻率檢測誘變物質的方法,由B.艾姆斯在1973年所首創,又稱艾姆斯測驗。測試對象是鼠傷寒沙門氏菌的幾個組氨酸缺陷型菌株,包括鹼基置換突變型和移碼突變型。在檢測系統中還包括大鼠的肝臟微粒體活化系統(S9),其中的酶能使一些前誘變劑轉變為誘變劑。雖然在這裡測試對象是細菌,而不是哺乳動物,但是由於這一檢測系統簡便易行,靈敏度較高,所以常用來作為誘變物質檢測初步篩選的短期測試系統,用這種方法已經對幾百種物質進行了測試,發現大約90%的致癌物質具有誘變作用。④中間宿主擴散盒法,為了能使回復突變法更接近於哺乳動物活體中的情況,有人把測試的細胞放在一種特製的小盒中,小盒的膜只允許溶液通過。把這種小盒埋藏在動物腹腔內,用待測物質處理動物,經過一定的時間后把小盒取出,測定小盒中被誘發回復突變的細胞數。
除了用來檢測基因突變的許多方法以外,還有許多用來檢測染色體畸變和姐妹染色單體互換的測試系統。當然對於藥物的致癌活性的最可靠的測定是哺乳動物體內致癌情況的檢測。但是利用微生物中誘發回復突變這一指標作為致癌物質的初步篩選,仍具有重要的實際意義(見毒理遺傳學)。
可以通過兩個途徑即鹼基結構類似物的參入和誘變劑或射線引起的化學變化來進行。
①類似物的參入5-溴尿嘧啶(BU)是胸腺嘧啶的結構類似物。它只是在第5位碳原子上以溴原子代替了胸腺嘧啶的甲基(─GH3),並且因此更易以烯醇式出現(圖2)。基因突變
大腸桿菌在含有BU的培養基中培養后,細菌的 DNA中的一部分胸腺嘧啶被BU所取代,並且最後在培養物中可以發現有少數突變型細菌出現,取代BU的量愈大則突變型愈多。突變型細菌在不含有BU的培養基中長久培養時,不改變它的突變型性狀,可是把突變型細菌在含有BU的培養基中培養后,又可以發現少數由於發生回復突變而出現的野生型細菌。BU的誘變作用可以表示。首先在DNA複製過程中酮式的BU代替了胸腺嘧啶T而使A:T鹼基對變為A:BU,在下一次DNA複製中烯醇式的BU*和鳥嘌呤G配對而出現G∶BU鹼基對,最後在又一次複製中鳥嘌呤G和胞嘧啶C配對而終於出現G:C鹼基對,完成了鹼基的置換。這裡BU所起的作用是促成這一置換,起促成作用的原因是由於嘧啶的 5位上溴原子代替了甲基后便較多地出現烯醇式的嘧啶。
同一理論還可以用來說明 BU是怎樣誘發 的置換突變或者突變型的回復突變(圖4)
2-氨基嘌呤等其他鹼基結構類似物同樣具有誘變作用。
②藥物或射線引起的化學變化亞硝酸能夠作用於腺嘌呤(A)的氨基而使它變為次黃嘌呤(HX);可以作用於胞嘧啶(c)而使它變為尿嘧啶(U)。這兩種氨基到酮基的變化帶來鹼基配對關係的改變,從而通過 DNA複製而造成A∶T→G∶C或者 G∶C→A∶T置換。
羥胺只和胞嘧啶發生專一性的反應,所以它幾乎只誘發置換G∶C→A∶T而不誘發A∶T→G∶C置換。此外,pH值低或高溫都可以促使DNA分子失去鹼基特別是嘌呤,導致鹼基置換。
紫外線的照射使 DNA分子上鄰接的鹼基形成二聚體,主要是胸腺嘧啶二聚體T-T。二聚體的形成使DNA雙鏈呈現不正常的構型(見DNA損傷修復),從而帶來致死效應或者導致基因突變,其中包括多種類型的鹼基置換。
誘發移碼突變的誘變劑種類較少,主要是吖啶類染料(圖6)。這些染料分子能夠嵌入DNA分子中,從而使DNA複製發生差錯而造成移碼突變。
利用重組DNA技術使DNA分子在指定位置上發生特定的變化,從而收到定向的誘變效果。例如將DNA分子用某一種限制性核酸內切酶處理,再用分解DNA單鏈的核酸酶S1處理,以去除兩個粘性末端的單鏈部分,然後用噬菌體T4連接酶將兩個平頭末端連接起來,這樣就可得到缺失了相應於這一限制性內切酶的識別位點的幾個核苷酸的突變型。相反地,如果在四種脫氧核苷三磷酸(dNTP)存在的情況下加入 DNA多聚酶Ⅰ,那麼進行互補合成的結果就得到多了相應幾個核苷酸的兩個平頭末端。在T4接連酶的處理下,便可以在同一位置上得到幾個核苷酸發生重複的突變型。
在指定的位置上也可以定向地誘發置換突變。誘變劑亞硫酸氫鈉能夠使胞嘧啶脫氨基而成為尿嘧啶,但是這種作用只限於 DNA單鏈上的胞嘧啶而對於雙鏈上的胞嘧啶則無效。用識別位點中包含一個胞嘧啶的限制性內切酶處理DNA分子,使粘性末端中的胞嘧啶得以暴露(例如HindⅢ的識別位點是,經限制酶HindⅢ處理后得到粘性末端,中間的這一胞嘧啶便暴露了)。經亞硫酸氫鈉處理后胞嘧啶(c)變為尿嘧啶(U)。通過DNA複製原來的鹼基對C∶G便轉變成為 T∶A。這樣一個指定位置的鹼基置換突變便被誘發。
還可以把人工合成的低聚核苷酸片段引入基因組中,以一定的方式改變某一基因等。
所謂自發突變是指未經誘變劑處理而出現的突變。從誘變機制的研究結果來看,自發突變的原因不外乎以下幾種。①背景輻射和環境誘變。短波輻射在宇宙中隨時都有,實驗說明輻射的誘變作用不存在閾效應,即任何微弱劑量的輻射都具有某種程度的誘變作用,因此自發突變中可能有一小部分是短波輻射所誘發的突變,有人估計果蠅的這部分突變約佔自發突變的 0.1%。此外,接觸環境中的誘變物質也是自發突變的一個原因。②生物自身所產生的誘變物質的作用。過氧化氫是一種誘變劑。在用過氧化氫作誘變處理時加入過氧化氫酶可以降低誘變作用,如果同時再加入氰化鉀(KCN)則誘變作用又重新提高。這是因為KCN是過氧化氫酶的抑製劑。另外又發現在未經誘變處理的細胞群體中加入KCN時,可以提高自發突變率,說明細胞自身所產生的過氧化氫是一部分自發突變的原因。在一些高等植物和微生物中曾經發現一些具有誘變作用的物質,在長久儲藏的洋蔥和煙草等種子中也曾經得到具有誘變作用的抽提物。③鹼基的異構互變效應。天然鹼基結構類似物5-溴尿嘧啶所以能誘發鹼基置換突變,是因為5位(圖2)上的溴原子促使BU較多地以烯醇式結構出現。在正常的情況下酮式和烯醇式之間的異構互變也以極低的頻率發生著,它必然同樣地造成一部分並不起源於環境因素的自發突變。此外,推測氨基和亞氨基之間的異構互變同樣是自發突變的一個原因。嚴格地講,這才是真正的自發突變。核苷酸還可以有其他形式的異構互變,它們同樣可能是自發突變的原因。
突變是一系列變化的結果。影響這一系列變化的任何一個環節的因素都會對於突變型的出現有一定的影響。
誘變劑接觸 DNA以前必須首先進入細胞,才能誘發突變。高等植物對於紫外線的誘變作用較不敏感的原因就是因為紫外線不易穿透它的細胞壁。化學藥品的滲透和細胞膜的結構有很大的關係。鼠傷寒沙門氏菌有一個改變細胞膜成分的突變型深度粗糙 (rfa),它使細胞膜對於許多藥物的滲透性增大,從而提高了細胞對許多化學誘變劑的敏感性。
細胞中的酶可以破壞進入細胞的誘變劑,從而減弱誘變效果。例如,過氧化氫酶可以減弱過氧化氫的誘變效果。一些沒有誘變作用的物質也可以因為細胞中的酶的活化作用而使該物質轉變成為誘變劑,這些物質稱為前誘變劑。例如陸蒽酮本身沒有誘變作用,但可以通過肝臟中的羥化酶的作用而轉變為誘變劑海蒽酮(圖7)。
誘變劑接觸DNA以後,能使DNA發生局部的損傷,這些損傷如果未經修復,便可阻礙 DNA的複製而造成細胞死亡。修復 DNA損傷的機制有兩類:一類稱為無誤修復,它使 DNA恢復原狀但不帶來突變;另一類稱為易誤修復或稱錯誤傾向修復,它使DNA複製繼續進行,但也常同時帶來基因突變。
細胞中有關 DNA損傷修復的酶活性的改變,可以改變細胞對於誘變劑的殺傷作用或誘變作用的反應。由於基因突變而使不論哪一種有關 DNA損傷修復的酶失活時,都必然導致細胞對於紫外線或其他誘變劑的殺傷作用變得更為敏感。可是就誘變結果來講,則要看這酶是涉及無誤修復,還是易誤修復。如果屬於前者,那麼有關的基因發生突變時將使突變更易發生,如果屬於後者,那麼有關的基因發生突變時將使突變更不易發生,因此這些突變型分別稱為增變基因和抗變基因。在大腸桿菌噬菌體T4中,基因43編碼 DNA多聚酶。基因43的突變型有兩種。一種是增變基因,它的 DNA多聚酶的核酸外切酶活性和多聚酶活性之比小於野生型的 DNA多聚酶;另一種是抗變基因,它的 DNA多聚酶的這兩種活性比大於野生型的 DNA多聚酶。在其他生物如大腸桿菌、酵母菌和一些真核生物中也曾發現增變基因。
1、鐮刀型細胞貧血症
(1)癥狀 紅細胞由正常的圓餅狀變成鐮刀型,導致紅細胞不能順利通過毛細血管聚集在一起,紅細胞破裂(溶血),造成貧血。
(2)病因 基因中的鹼基替換。直接原因:血紅蛋白分子結構的改變 根本原因:控制血紅蛋白分子合成的基因結構的改變。
2、基因突變
概念:DNA分子中發生鹼基對的替換、增添和缺失,而引起的基因結構的改變。
外界因素
①溫度,基因突變包括一系列生物化學變化,所以溫度對於基因突變有一定的影響。在大腸桿菌中,組氨酸缺陷型(his-)在15℃到37℃範圍內溫度每升高 10℃自發回復突變率提高1~1.5倍,在0℃時不發生自發突變。果蠅的致死突變的溫度係數也在這範圍內。在微生物和果蠅中,較短時間的溫度改變,特別是不適宜於生存的較高溫度的處理,都可以誘發突變;在果蠅中還有-6℃低溫處理誘發突變的報道。②培養基成分,SOS是一種經誘導后才出現的易誤修復機制。和誘導酶的合成一樣,蛋白質合成是使細菌細胞中出現SOS機制的必要因素,所以培養基中一切影響蛋白質合成的因素都會影響基因突變。③抗變劑和助變劑,能夠促進另一誘變劑的作用的物質稱為助變劑。例如,色氨酸燒焦后產生兩種誘變劑和助變劑。
已經知道亞硝基胍(NTC)是一種高效誘變劑。在一定條件下,NTG的誘變效果能被氯化鈷和活體紅細胞中的含硫化合物減低,說明氯化鈷和紅細胞中存在著的某種物質具有抗變作用。這些能夠降低自發或誘發突變率的物質稱為抗變劑。此外,某些多肽如白肽素等也都被證明是抗變劑。

基因突變
加州大學舊金山分校的生物化學和生物物理學教授Elizabeth Blackburn(伊麗莎白)說:“這就象毒針一樣,你只需要加一點點就可以得到顯著的效果。”
這種突變的目標是一個在癌細胞中高度活躍的酶——端粒酶,該酶在細胞複製的消耗過程中幫助維持染色體結構。
該突變使用端粒酶來破壞迅速擴增的癌細胞——Blackburn將這個策略比作柔道,雙方利用對手的力量來擊敗對方。
在這項研究中,科學家在該酶的遺傳密碼中插入了一個由RNA構成的小突變。突變的RNA阻斷了端粒酶將RNA反轉錄為DNA,以重建細胞複製過程中丟失的染色體部分的正常活性。
Blackburn說:“癌細胞是著名的對抗自殺信號的細胞類型,這是之所以癌細胞如此可怕的原因之一。擁有這麼少量的端粒酶能夠發揮如此有效的作用相當令人吃驚。”
研究中,低水平的突變RNA大大降低了乳腺癌和前列腺癌細胞的生長速度,並且使更多的細胞死亡。
這種突變在導入突變酶的活體小鼠中使得乳腺癌腫瘤減小。
基因突變
的研究可能發現來自人體的癌細胞比研究中使用的實驗室培養的細胞對突變酶要更為敏感。
Blackburn和同事們完成的這項研究發表在最新一期的Proceedingsof the National Academy of Sciences上。科學家們一直在研究通過破壞端粒酶活性來治療癌症的幾種方法,但是加州大學進行的這項研究提供了新的治療。國家健康研究所的Richard Hodes說:“在被研究的幾個供選擇方法中,端粒酶突變作為直接影響腫瘤細胞治療癌症的方法具有清晰的理論優勢。”
基本信息
- 中文名
- 基因突變
- 外文名
- genic mutation
- 提出時間
- 1959年
- 影響
- 同義突變、錯義突變
- 提出者
- E.佛里茲
- 應用學科
- 生物
- 適用領域
- 醫學