等位基因
控制相對性狀的基因
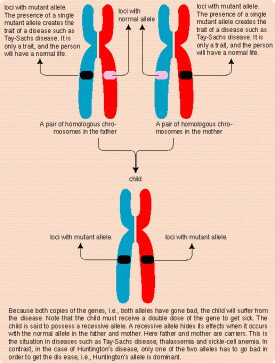
等位基因(allele),是指位於一對同源染色體相同位置上控制同一性狀不同形態的基因。
註釋:同源染色體是在二倍體生物細胞中,形態、結構基本相同的染色體,並在減數第一次分裂(參考減數分裂)的四分體時期中彼此聯會(若是三倍體及其他奇數倍體生物細胞,聯會時會發生紊亂),最後分開到不同的生殖細胞(即精子、卵細胞)的一對染色體,在這一對染色體其中的一條來自母方,另一條來自父方。
我們先了解一些基本概念:生物的形態、結構、生理特徵稱為性狀,比如人的眼瞼形態就是一種性狀,這種性狀有不同的表現形式:雙重瞼(俗稱雙眼皮)、單重瞼( superiorepiblepharon上瞼贅皮,俗稱單眼皮),其中單重瞼為隱性,雙重瞼為顯性。我們把它們稱為相對性狀(其概念是同種生物同一性狀的不同表現類型)。性狀又是由基因控制的,控制顯性性狀的為顯性基因 (用大寫字母,如A),控制隱性性狀的為隱性基因(用小寫字母,如a),基因在體細胞中成對存在,所以一個個體的基因型就有:AA,Aa,aa,不過也有發生了染色體變異導致有多個基因。A和a就可以表示一對等位基因。它們的定義為:同源染色體的相同位置上,控制相對性狀的一對基因。
染色體上的等位基因
以上來自《大英百科全書中文版》 allele 等位基因條。國內個別教材上D和D,d和d不是等位基因的說法是錯誤的。錯誤在於,沒有理解所謂等位基因是多個基因中的一個,而是從中文字面上望文生義,想當然的認為只有兩個基因,才能相互之間叫做等位基因,所以才會有D和D是不是等位基因的問題。而且更進一步認為,只有D和d才能相互之間叫做等位基因,這就更不對了。(註:國內高中課本必修二課本將等位基因定義為:控制一對相對性狀的基因。而由於D與D不是控制一對相對性狀的基因所以不能稱作等位基因.)
在一個個體里,某個基因的基因型是由該基因所擁有的一組等位基因所決定。例如,二倍體生物,也就是每條染色體都有兩套的生物,兩個等位基因決定了該基因的基因型。
位於一對同源染色體的相同位置上控制某一性狀的不同形態的基因。不同的等位基因產生例如發色或血型等遺傳特徵的變化。靠等位基因控制相對性狀的顯隱性關係及遺傳效應,可將等位基因區分為不同的類別。在個體中,等位基因的某個形式(顯性的)可以比其他形式(隱性的)表達得多。
位於一對同源染色體的相同位置上控制某一性狀的不同形態的基因。不同的等位基因產生例如發色或血型等遺傳特徵的變化。等位基因控制相對性狀的顯隱性關係及遺傳效應,可將等位基因區分為不同的類別。在個體中,等位基因的某個形式(顯性的)可以比其他形式(隱性的)表達得多。等位基因(gene)是同一基因的另外“版本”。例如,控制捲舌運動的基因不止一個“版本”,這就解釋了為什麼一些人能夠捲舌,而一些人卻不能。有缺陷的基因版本與某些疾病有關,如囊性纖維化。值得注意的是,每個染色體(chromosome)都有一對“複製本”,一個來自父親,一個來自母親。這樣,我們的大約3萬個基因中的每一個都有兩個“複製本”。這兩個複製本可能相同(相同等位基因allele),也可能不同。下圖顯示的是一對染色體,上面的基因用不同顏色表示。在細胞分裂過程中,染色體的外觀就是如此。如果比較兩個染色體(男性與女性)上的相同部位的基因帶,你會看到一些基因帶是相同的,說明這兩個等位基因是相同的;但有些基因帶卻不同,說明這兩個“版本”(即等位基因)不同。
(pseudoalleles):表型效應相似,功能密切相關,在染色體上的位置又緊密連鎖的基因。它們象是等位基因,而實際不是等位基因。
傳統的基因概念由於擬等位基因現象的發現而更趨複雜。摩根學派在其早期的發現中特別使他們感到奇怪的是相鄰的基因一般似乎在功能上彼此無關,各行其是。影響眼睛顏色、翅脈形成、剛毛形成、體免等等的基因都可能彼此相鄰而處。具有非常相似效應的“基因”一般都僅僅不過是單個基因的等位基因。如果基因是交換單位,那就絕不會發生等位基因之間的重組現象。事實上摩根的學生在早期(1913;1916)試圖在白眼基因位置發現等位基因的交換之所以都告失敗,後來才知道主要是由於試驗樣品少。然而自從斯特體範特(1925)提出棒眼基因重複的不均等交換學說以及布里奇斯(1936)根據唾液腺染色體所提供的證據支持這學說之尼,試圖再一次在彷彿是等位基因之間進行重組的時機已經成熟。Oliver(1940)首先取得成功,在普通果蠅的菱形基因位置上發現了等位基因不均等交換的證據。兩個不同等位基因(Izg/Izp)被標誌基因拚合在一起的雜合子以0.2%左右的頻率回復到野生型。標誌基因的重組證明發生了“等位基因”之間的交換。
非常靠近的基因之間的交換隻能在極其大量的試驗樣品中才能觀察到,由於它們的正常行為好像是等位基因,因此稱為擬等位基因(Lewis,967)。它們不僅在功能上和真正的等位基因很相似,而且在轉位(transposition)后能產生突變體表現型。它們不僅存在於果蠅中,而且在玉米中也已發現,特別在某些微生物中發現的頻率相當高。分子遺傳學對這個問題曾有很多解釋,然而由於目前對真核生物的基因調節還知之不多,所以還無法充分了解。
位置效應的發現產生了深刻影響。杜布贊斯基在一篇評論性文章中曾對此作出下面的結論;“一個染色體不單是基因的機械性聚合體,而且是更高結構層次的單位……染色體的性質由作為其結構單位的基因的性質來決定;然而染色體是一個合諧的系統,它不僅反映了生物的歷史,它本身也是這歷史的一個決定因素”(Dobzhaansky,1936:382)。
有些人並不滿足於這種對基因的“串珠概念”的溫和修正。自從孟德爾主義興起之初就有一些生物學家(例如Riddle和Chiid)援引了看來是足夠份量的證據反對基因的顆粒學說。位置效應正好對他們有利。Goldschmidt(1938;1955)這時變成了他們的最雄辯的代言人。他提出一個“現代的基因學說”(1955:186)來代替(基因的)顆粒學說。按照他的這一新學說並沒有定位的基因而只有“在染色體的一定片段上的一定分子模式,這模式的任何變化(最廣義的位置效應)就改變了染色體組成部分的作用從而表現為突變體。”染色體作為一個整體是一個分子“場”,習慣上所謂的基因是這個場的分立的或甚至是重疊的區域;突變是染色體場的重新組合。這種場論和遺傳學的大量事實相矛盾因而未被承認,但是像Goldschmidt這樣一位經驗豐富的知名遺傳學家竟然如此嚴肅地提出這個理論這件事實就表明基因學說還是多麼不鞏固。從1930年代到1950年代所發表的許多理論性文章也反映了這一點(Demerec,1938,1955;Muller,1945;Stadler,1954)。
若同源染色體上同一位置上的等位基因的數目在兩個以上,就稱為復等位基因(multiple allelism)。任何一個二倍體個體只存在復等位基因中的二個不同的等位基因。
在完全顯性中,顯性基因中純合子和雜合子的表型相同。在不完顯性中雜合子的表型是顯性和隱性兩種純合子的中間狀態。這是由於雜合子中的一個基因無功能,而另一個基因存在劑量效應所致。完全顯性中雜合體的表型是兼有顯隱兩種純合子的表型。此是由於雜合子中一對等位基因都得到表達所致。
比如決定人類ABO血型系統四種血型的基因IA、IB、i,每個人只能有這三個等位基因中的任意兩個。
1910年美國遺傳學家摩爾根(T. H. Morgan)證明基因位於染色體上,並把位於同一條染色體上的基因稱為連鎖群。大多數真核生物的體細胞是二倍體細胞,細胞里的染色體是成對存在的,二者互為同源染色體;而生殖細胞里每種染色體都只有一條,所以是單倍體細胞。二倍體細胞每個基因也是成對存在的,每一對基因分別位於來自雙親的染色體的同一位置上,這個位置稱為基因座。一對同源染色體同一基因座上的一對基因稱為一對等位基因。例如,豌豆的高莖基因與矮莖基因互為等位基因。一個二倍體生物如果具有一對不同的等位基因,則這種生物為該基因的雜合子,反之則為純合子。若雜合子的一對等位基因中只有一個能表達出性狀,另一個不表達,則前者稱顯性基因,後者稱隱性基因。如果一對等位基因同時表達,則稱為共顯性。
對個體而言,一對同源染色體的一個基因座上只有一對等位基因。但在一個生物群體中,一個基因座上的等位基因多於兩種時,稱為復等位基因。例如決定人類ABO血型系統的等位基因有三種,分別為IA、IB和i。就每個人而言,只可能具有這3種復等位基因中的1種或2種,從而表現出特定的血型。在這裡,A和B對i而言是顯性,IA和IB是共顯性,i是隱性。
重組等位基因形成的
等位基因(allele):位於一對同源染色體的相同位置上控制某一性狀的不同形態的基因。不同的等位基因產生例如發色或血型等遺傳特徵的變化。等位基因控制相對性狀的顯隱性關係及遺傳效應,可將等位基因區分為不同的類別。在個體中,等位基因的某個形式(顯性的)可以比其他形式(隱性的)表達得多。
例如,人類RH血型基因的座位是在1號染色體短臂的3區5帶,位於兩條1號染色體相同座位的Rh和RH就是一對等位基因。
在一個群體內,同源染色體的某個相同座位上的等位基因超過2個以上時,就稱作復等位基因。例如,人類 ABO 血型基因座位是在9號染色體長臂的末端,在這個座位上的等位基因,就人類來說,有IA、IB、i三個基因,因此人類的 ABO血型是由3個復等位基因決定的。但就一個具體人類來說,決定 ABO 血型的一對等位基因,是A、B、O三個基因中的兩個,即IAIA、IBIB、IAIB、ii、IAi、IBi當一個生物體帶有一對完全相同的等位基因時,則該生物體就該基因而言是純合的(homozygous)或可稱純種(true-breeding);反之,如果一對等位基因不相同,則該生物體是雜合的(heterozygous)或可稱雜種(hybrid)。等位基因各自編碼蛋白質產物,決定某一性狀,並可因突變而失去功能。
等位基因之間存在相互作用。當一個等位基因決定生物性狀的作用強於另一等位基因並使生物只表現出其自身的性狀時,就出現了顯隱性關係。作用強的是顯性,作用被掩蓋而不能表現的為隱性。一對呈顯隱性關係的等位基因,顯性完全掩蓋隱性的是完全顯性(complete dominance),兩者相互作用而出現了介於兩者之間的中間性狀,如紅花基因和白花基因的雜合體的花是粉紅色,這是不完全顯性(incomplete dominance)。有些情況下,一對等位基因的作用相等,互不相讓,雜合子就表現出兩個等位基因各自決定的性狀,這稱為共顯性(codominance)。1946年,談家楨在亞洲異色瓢蟲(Hormonia axynidis)鞘翅的色斑遺傳現象中發現的嵌鑲顯性(mo—saic dominance)就是共顯性的一個特殊例子。
亞洲異色瓢蟲鞘翅的底色為黃色,底色上有各種形狀的黑色斑點,形成不同的圖案。子代瓢蟲的鞘翅能同時顯現出父本和母本的黑色斑點,相同位置上的顏色互相重疊,黑色掩蓋了黃色。嵌鑲顯性是由復等位基因控制的。
野生型(wild type)用來描述自然界中常見的基因型和表現型。野生型等位基因都產生有功能的蛋白質。突變型等位基因最常見的是喪失功能型(loss-of-function),絕大多數產生改變了的蛋白質,極少數根本不產生蛋白質。所以,野生型對突變型而言是顯性。但是,如果突變型等位基因是獲得功能型(gain-of-function),產生的蛋白質賦予生物體以新的性狀,此時突變型等位基因則為顯性。對一個二倍體細胞而言,當一個等位基因的功能已足夠使某個性狀表現時,這個等位基因就表現為完全顯性;而當二倍體細胞的某性狀表現對等位基因的功能有數量上的要求時,例如,需要等位基因的兩份活性產物,則雜合子就表現為不完全顯性。
一對不同的等位基因各有自己特定的產物和表型,雜合子同時表現出雙親的特性,則是共顯性。
非等位基因之間也存在相互作用。位於同一染色體的不同基因座,或位於不同染色體上的非等位基因,都可能影響到同一性狀。例如,某些性狀只有同時存在若干個非等位基因時才會出現,當其中任何一個非等位基因發生改變時,都會導致產生同一種突變性狀。這些非等位基因稱為互補基因(complementary gene)。又如,有些基因本身沒有可觀察到的表型效應,但可以抑制其他非等位基因的活性,這就是抑制基因(inhibitor)。
上位效應(epistasis)則指一對基因可以掩蓋另一對非等位基因的顯性效應的現象,這是非等位基因之間的掩蓋作用,也可以稱為異位效應。非等位基因之間的相互作用實質上是基因表達的順式調控或反式調控的結果。