同源染色體
同源染色體
同源染色體是在二倍體生物細胞中,形態、結構基本相同的染色體,並在減數第一次分裂(參考減數分裂)的四分體時期中彼此聯會(若是三倍體及其他奇數倍體生物細胞,聯會時會發生紊亂),最後分開到不同的生殖細胞(即精子、卵細胞)的一對染色體,在這一對染色體其中的一個來自母方,另一個來自父方。
同源染色體
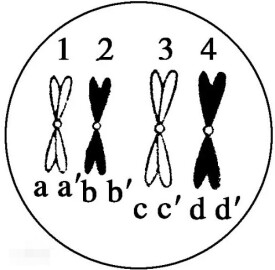
正常細胞中,一對同源染色體若表示為由兩條染色體A、B組成,在減數分裂中變成“AA`”組和“BB`”組,兩組聯會形成四分體;有絲分裂中變成“AA`”組和“BB`”組,但兩組不聯會,在有絲分裂中期染色體的著絲點整齊排列在赤道板上,著絲點分開后染色體平均分配到兩個子細胞。
在生物體的有性生殖過程中,有性生殖細胞是通過細胞分裂的一種——減數分裂形成的。在減數分裂的分裂間期,精原細胞的體積略微增大,染色體進行複製,成為初級精母細胞。複製后的每條染色體都含有兩條姐妹染色體,這兩條姐妹染色單體並列在一起,由同一個著絲點連接著。分裂期開始后不久,初級精母細胞中原來分散存在的染色體進行配對。而在減數第二次分裂過程中不存在同源染色體。
區分同源染色體與姐妹染色單體:姐妹染色單體是由一個著絲點連著的并行的兩條染色單體,是在細胞分裂的間期由同一條染色體經複製后形成的——由一條染色體複製形成的兩條子染色體不是同源染色體,因為它們儘管形狀大小相同,但它們並非一條來自父方、一條來自母方。
不僅只有減數分裂有同源染色體的存在,有絲分裂也有。所有的只要有一對染色體存在的都會存在。
在減數分裂中的功能
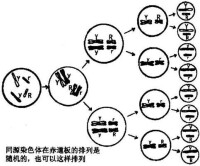
減數分裂(Meiosis)進行兩次細胞分裂,產生四個單倍體子細胞,每個子細胞含有親體細胞的一半染色體。它首先通過分離減數分裂I期中的同源染色體,再通過分離減數分裂II期中的姐妹染色單體,將生殖細胞中的染色體數量減少一半。減數分裂I期的過程通常比減數分裂II期長,因為染色質複製需要更多的時間,並且同源染色體在分裂I期通過配對和染色體聯會適當定位和分離也需要時間。減數分裂過程的遺傳重組(通過隨機分離)和交會產生的每個子細胞,均含有來自母體和父體編碼基因的不同組合。這種基因重組允許引入新的等位基因配對和遺傳變異。生物間的遺傳變異通過提供更廣泛的自然選擇的遺傳特性來促使種群更加穩定。因此,減數分裂不僅是保證生物種染色體數目穩定的機制,同且也是物種適應環境變化不斷進化的機制。
1 前期I
減數分裂I期的前期I中,DNA已經經歷了複製,因此,每個染色體由兩個相同的染色單體組成(姐妹染色體),這些染色體由一個共同的著絲粒連接。在前期I的合子階段,同源染色體相互配對。這種配對通過聯會過程發生,其中聯會複合體 被組裝並沿著它們的長度方向與同源染色體連接。發生在同源染色體之間內聚交聯,有助於它們抵抗同源染色體的被拉開直到減數分裂後期。遺傳交叉是一種重組,發生在前期I的粗線期。另外經常發生的另一種類型的重組是合成依賴性鏈退火重組(SDSA)。 SDSA重組涉及配對同源染色單體之間的信息交換,但不涉及物理交換。 SDSA重組也不會導致交叉。在交叉過程中的基因的交換是通過染色體同源部分的斷裂和結合來實現的。交換髮生在稱為交叉站點的場所。一旦交叉發生,該交叉站點將於同源染色體結合,並伴隨其後的染色體分離過程。無論是非交換還是交換類型的重組都能起到修復DNA損傷作用,特別是雙鏈斷裂的修復。在前期I的雙線期階段,聯會複合體分解,在此之前同源染色體分離,但姐妹染色單體仍通過著絲粒保持不分離。
2 中期I
在減數分裂I的中期I中,同源染色體對(也稱為二價體或四分體)沿著中期板隨機排列。這種隨機排列也是細胞引入遺傳變異的另一種方式。此時,從紡錘體主軸桿兩端發出的減數分裂紡錘體通過著絲點與每個同源物(每對姐妹染色單體)相連。
3 後期I
在減數分裂I的後期I中,同源染色體彼此分開。同源物被分離酶切割后,會釋放出將同源染色體臂保持在一起的粘連蛋白。這使得交叉站點得到釋放,同源物移動到細胞的相反極。同源染色體被隨機分離到兩個子細胞中,這兩個子細胞將經歷減數分裂II以產生四個單倍體子代生殖細胞。
4 減數分裂II
在減數分裂I中同源染色體的四分體分離之後,姐妹染色單體也進行了分離。兩個單倍體(因為染色體數目減少了一半)子細胞在減數分裂II中進行另一次細胞分裂,但不進行另一輪染色體複製。兩個子細胞中的姐妹染色單體在後期II被核紡絲纖維拉開,產生四個單倍體子細胞。
在有絲分裂中的功能
同源染色體在有絲分裂中的功能與減數分裂中的功能不相同。在每個細胞經歷有絲分裂之前,親體細胞中的染色體會自身複製。但細胞內的同源染色體通常不會配對也不進行基因重組。相反,複製子或姐妹染色單體將沿著中期板排列,然後以與減數分裂II相同的方式分離,即通過核有絲分裂紡錘體在它們的著絲粒處被拉開。即使在有絲分裂期間姐妹染色單體之間確實發生了交叉,也不會產生任何新的重組基因型。
在體細胞中 的功能
在大多數情況下,同源配對發生在生殖細胞中,但體細胞中也有發生。例如,人類體細胞具有非常嚴格調節的同源配對(分離成染色體疆域,在發育信號控制下在特定基因位點配對。其他物種(特別是果蠅)同源配對更頻繁。在21世紀早期的高通量篩選闡明了體細胞中同源配對的各種功能。
為什麼同源染色體是一條來自母方一條來自父方?
答:因為在減數分裂以後形成的配子(生殖細胞)中沒有同源染色體,所以在受精作用后,精子的細胞核和卵子的細胞核融合,進而染色體數加倍,在這以後就又有了同源染色體,其中的一條來自母方而另一條來自父方。
減數分裂(Meiosis)的特點是DNA複製一次,而細胞連續分裂兩次,形成單倍體的精子和卵子(圖13-12),通過受精作用又恢復二倍體,減數分裂過程中同源染色體間發生交換,使配子的遺傳多樣化,增加了後代的適應性,因此減數分裂不僅是保證生物種染色體數目穩定的機制,同且也是物種適應環境變化不斷進化的機制。減數分裂可分為3種主要類型:
也叫終端減數分裂(terminal meiosis),其特點是減數分裂和配子的發生緊密聯繫在一起,在雄性脊椎動物中,一個精母細胞經過減數分裂形成4個精細胞,後者在經過一系列的變態發育,形成成熟的精子。在雌性脊椎動物中,一個卵母細胞經過減數分裂形成1個卵細胞和2-3個極體。
也叫中間減數分裂(intermediate meiosis),見於植物和某些藻類。其特點是減數分裂和配子發生沒有直接的關係,減數分裂的結果是形成單倍體的配子體(小孢子和大孢子)。小孢子再經過兩次有絲分裂形成包含一個營養核和兩個雄配子(精子)的成熟花粉(雄配子體),大孢子經過三次有絲分裂形成胚囊(雌配子體),內含一個卵核、兩個極核、3個反足細胞和兩個助細胞。
也叫初始減數分裂(initial meiosis),僅見於真菌和某些原核生物,減數分裂發生於合子形成之後,形成單倍體的孢子,孢子通過有絲分裂產生新的單倍體後代。
此外某些生物還具有體細胞減數分裂(somatic meiosis)現象,如在蚊子幼蟲的腸道中,有一些由核內有絲分裂形成的多倍體細胞(可高達32X),在蛹期又通過減數分裂降低了染色體倍性,增加了細胞數目。
減數分裂由緊密連接的兩次分裂構成。通常減數分裂I分離的是同源染色體,所以稱為異型分裂(heterotypic division)或減數分裂(reductional division)。減數分裂II分離的是姊妹染色體,類似於有絲分裂,所以稱為同型分裂(homotypic division)或均等分裂(equational division)。和有絲分裂一樣為了描述方便將減數分裂分為幾個期和亞期。
有絲分裂細胞在進入減數分裂之前要經過一個較長的間期,稱前減數分裂間期(premeiotic interphase)或前減數分裂期(premeiosis)。
前減數分裂期也可分為G1期、S期和G2期,在G1期和S期把麝香百合的花粉每細胞在體外培養,則發現細胞進行有絲分裂,將G2晚期的細胞在體外培養則向減數分裂進行,說明G2期是有絲分裂向減數分裂轉化的關鍵時期。
和有絲分裂不同的是,DNA不僅在S期合成,而且也在前期合成一小部分。D. E. Wimber和W. Prensky(1963)認為合線期-粗線期合成大約2%的DNA。Y. Hotta等人(1966)在百合屬(Lilium)和延齡草屬(Trillium)中發現,粗線期合成大約0.3%的DNA。稱為合線期DNA(zyg-DNA)或粗線期DNA(P-DNA)。這些DNA的合成可能與聯會複合體的形成有關。
一、減數分裂I
1.前期I
減數分裂的特殊過程主要發生在前期I,通常人為劃分為5個時期:①細線期(leptotene)、②合線期(zygotene)、③粗線期(pachytene)、④雙線期(diplotene)、⑤終變期(diakinesis)。必須注意的是這5個階段本身是連續的,它們之間並沒有截然的界限。
1)細線期:
染色體呈細線狀,具有念珠狀的染色粒。持續時間最長,占減數分裂周期的40%。細線期雖然染色體已經複製,但光鏡下分辨不出兩條染色單體。由於染色體細線交織在一起,偏向核的一方,所以又稱為凝線期(synizesis),在有些物種中表現為染色體細線一端在核膜的一側集中,另一端放射狀伸出,形似花束,稱為花束期(bouquet stage)。
2)合線期:持續時間較長,占減數分裂周期的20%。亦稱偶線期,是同源染色體配對的時期,這種配對稱為聯會(synapsis)。這一時期同源染色體間形成聯會複合體(synaptonemal complex,SC)。在光鏡下可以看到兩條結合在一起的染色體,稱為二價體(bivalent)。每一對同源染色體都經過複製,含四個染色單體,所以又稱為四分體(tetrad)。
3)粗線期:持續時間長達數天,此時染色體變短,結合緊密,在光鏡下只在局部可以區分同源染色體,這一時期同源染色體的非姊妹染色單體之間發生交換的時期。在果蠅粗線期SC上具有與SC寬度相近的電子緻密球狀小體,稱為重組節,與DNA的重組有關。
4)雙線期:聯會的同源染色體相互排斥、開始分離,但在交叉點(chiasma)上還保持著聯繫。雙線期染色體進一步縮短,在電鏡下已看不到聯會複合體。
交叉的數目和位置在每個二價體上並非是固定的,而隨著時間推移,向端部移動,這種移動現象稱為端化(terminalization),端化過程一直進行到中期。
植物細胞雙線期一般較短,但在許多動物中雙線期停留的時間非常長,人的卵母細胞在五個月胎兒中已達雙線期,而一直到排卵都停在雙線期,排卵年齡大約在12-50歲之間。成熟的卵細胞直到受精后,才迅速完成兩次分裂,形成單倍體的卵核。
在魚類、兩棲類、爬行類、鳥類以及無脊椎動物的昆蟲中,雙線期的二價體解螺旋而形成燈刷染色體,這一時期是卵黃積累的時期。
5)終變期:二價體顯著變短,並向核周邊移動,在核內均勻散開。所以是觀察染色體的良好時期。
由於交叉端化過程的進一步發展,故交叉數目減少,通常只有一至二個交叉。終變期二價體的形狀表現出多樣性,如V形、O形等。
核仁此時開始消失,核被膜崩解,但有的植物,如玉米,在終變期核仁仍然很顯著。
2.中期I
核仁消失,核被膜崩解,標誌進入中期I,中期I的主要特點是染色體排列在赤道面上。每個二價體有4個著絲粒、姊妹染色單體的著絲粒定向於紡錘體的同一極,故稱聯合定向(co-orientation)。
3.後期I
二價體中的兩條同源染色體分開,分別向兩極移動。由於相互分離的是同源染色體,所以染色體數目減半。但每個子細胞的DNA含量仍為2C。同源染色體隨機分向兩極,使母本和父本染色體重所組合,產生基因組的變異。如人類染色體是23對,染色體組合的方式有2^23個(不包括交換),因此除同卵孿生外,幾乎不可能得到遺傳上等同的後代。
4.末期I
染色體到達兩極后,解旋為細絲狀、核膜重建、核仁形成,同時進行胞質分裂。
5.減數分裂間期
在減數分裂I和II之間的間期很短,不進行DNA的合成,有些生物沒有間期,而由末期I直接轉為前期II。
二、減數分裂II
可分為前、中、后、末四個四期,與有絲分裂相似。
通過減數分裂一個精母細胞形成4個精子。
而一個卵母細胞形成一個卵子及2-3個極體。
聯會複合體(synaptonemal complex,SC)是減數分裂合線期兩條同源染色體之間形成的一種結構,它與染色體的配對,交換和分離密切相關。
SC是同源染色體間形成的梯子樣的結構。在電鏡下觀察,兩側是約40nm的側生組分(lateral element),電子密度很高,兩側之間為寬約100nm的中間區(intermediate space),在電鏡下是明亮區,在中間區的中央為中央組分(central element),寬約30nm。側生組分與中央組分之間有橫向排列的粗約7~10nm的SC纖維,使SC外觀呈梯子狀。
長期以來人們認為SC將同源染色體組織在一起,使伸入SC的DNA之間產生重組,但實驗證明不僅SC的形成晚於基因重組的啟動,而且基因突變不能形成SC的酵母中,同源染色體間照樣可以發生交換。一般認為它與同源染色體間交換的完成有關。
在磷鎢酸染色的SC中央,還可以看到呈圓形或橢圓形的重組節(recombination nodules,RNs),RNs是同源染色體發生交叉的部位,RNs上有基因交換所需要的酶。
從形態學來看,SC形成合線期,成熟於粗線期,並存在數天,消失於雙線期。聯會複合體的形成與合線期DNA(Zyg-DNA)有關,在細線期或合線期加入DNA合成抑製劑,則抑制SC的形成。