遺傳學
研究生物遺傳與變異的科學
遺傳學(Genetics)——自然科學領域中探究生物遺傳和變異規律的科學。
研究基因的結構、功能及其變異、傳遞和表達規律的學科(全國科學技術名詞審定委員會審定·遺傳學名詞。北京:科學出版社,2006年3月)。
遺傳學(Genetics)——研究生物的遺傳與變異的科學,研究基因的結構、功能及其變異、傳遞和表達規律的學科。遺傳學中的親子概念不限於父母子女或一個家族,還可以延伸到包括許多家族的群體,這是群體遺傳學的研究對象。遺傳學中的親子概念還可以以細胞為單位,離體培養的細胞可以保持個體的一些遺傳特性,如某些酶的有無等。對離體培養細胞的遺傳學研究屬於體細胞遺傳學。遺傳學中的親子概念還可以擴充到脫氧核糖核酸(也就是DNA)的複製甚至mRNA的轉錄,這些是分子遺傳學研究的課題。
一。基因
基因存在於生物的細胞核,染色體,DNA中。染色體是成對出現的,所以DNA,基因都是成對出現的。正常情況下,人體內有23對染色體,46個DNA分子,無數個基因(一個DNA中有無數個基因)。
二。遺傳學研究範圍
染色體

遺傳學
由一個受精卵產生的免疫活性細胞能夠分別產生各種不同的抗體球蛋白,這也是遺傳學的一個課題,它的研究屬於免疫遺傳學。
從噬菌體到人,生物界有基本一致的遺傳和變異規律,所以遺傳學原則上不以研究的生物對象劃分學科分支。人類遺傳學的劃分是因為研究人的遺傳學與人類的幸福密切相關,而系譜分析和雙生兒法等又幾乎只限於人類的遺傳學研究。
微生物遺傳學的劃分是因為微生物與高等動植物的體制很不相同,因而必須採用特殊方法進行研究。此外,還有因生產意義而出現的以某一類或某一種生物命名的分支學科,如家禽遺傳學、棉花遺傳學、水稻遺傳學等。
更多的遺傳學分支學科是按照所研究的問題來劃分的。例如,細胞遺傳學是細胞學和遺傳學的結合;發生遺傳學所研究的是個體發育的遺傳控制;行為遺傳學研究的是行為的遺傳基礎;免疫遺傳學研究的是免疫機制的遺傳基礎;輻射遺傳學專門研究輻射的遺傳學效應;藥物遺傳學則專門研究人對藥物反應的遺傳規律和物質基礎,等等。
從群體角度進行遺傳學研究的學科有群體遺傳學、生態遺傳學、數量遺傳學、進化遺傳學等。這些學科之間關係緊密,界線較難劃分。群體遺傳學常用數學方法研究群體中的基因的動態,研究基因突變、自然選擇、群體大小、交配體制、遷移和漂變等因素對群體中的基因頻率和基因平衡的影響;生態遺傳學研究的是生物與生物,以及生物與環境相互適應或影響的遺傳學基礎,常把野外工作和實驗室工作結合起來研究多態現象、擬態等,藉以驗證群體遺傳學研究中得來的結論;進化遺傳學的研究內容包括生命起源、遺傳物質、遺傳密碼和遺傳機構的演變以及物種形成的遺傳基礎等。物種形成的研究也和群體遺傳學、生態遺傳學有密切的關係。
從應用角度看,醫學遺傳學是人類遺傳學的分支學科,它研究遺傳性疾病的遺傳規律和本質;臨床遺傳學則研究遺傳病的診斷和預防;優生學則是遺傳學原理在改良人類遺傳素質中的應用。生統遺傳學或數量遺傳學的主要研究對象是數量性狀,而農作物和家畜的經濟性狀多半是數量性狀,因此它們是動植物育種的理論基礎。
雜交是遺傳學研究的最常用的手段之一,所以生活周期的長短和體形的大小是選擇遺傳學研究材料常要考慮的因素。昆蟲中的果蠅、哺乳動物中的小鼠和種子植物中的擬南芥,便是由於生活周期短和體形小而常被用作遺傳學研究的材料。大腸桿菌和它的噬菌體更是分子遺傳學研究中的常用材料。
生物化學方法幾乎為任何遺傳學分支學科的研究所普遍採用,更為分子遺傳學所必需。分子遺傳學中的重組DNA技術或遺傳工程技術已逐漸成為遺傳學研究中的有力工具。

孟德爾
許多人在這些活動的基礎上力圖闡明親代和雜交子代的性狀之間的遺傳規律都未獲成功。直到1866年奧地利學者孟德爾根據他的豌豆雜交實驗結果發表了《植物雜交試驗》的論文,揭示了現在稱為孟德爾定律的遺傳規律,才奠定了遺傳學的基礎。
孟德爾的工作結果直到20世紀初才受到重視。19世紀末葉在生物學中,關於細胞分裂、染色體行為和受精過程等方面的研究和對於遺傳物質的認識,這兩個方面的成就促進了遺傳學的發展。
從1875~1884的幾年中德國解剖學家和細胞學家弗萊明在動物中,德國植物學家和細胞學家施特拉斯布格在植物中分別發現了有絲分裂、減數分裂、染色體的縱向分裂以及分裂后的趨向兩極的行為;比利時動物學家貝內登還觀察到馬副蛔蟲的每一個身體細胞中含有等數的染色體;德國動物學家赫特維希在動物中,施特拉斯布格在植物中分別發現受精現象;這些發現都為遺傳的染色體學說奠定了基礎。美國動物學家和細胞學家威爾遜在 1896年發表的《發育和遺傳中的細胞》一書總結了這一時期的發現。
關於遺傳的物質基礎歷來有所臆測。例如1864年英國哲學家斯賓塞稱之為活粒;1868年英國生物學家達爾文稱之為微芽; 1884年瑞士植物學家內格利稱之為異胞質;1889年荷蘭學者德弗里斯稱之為泛生子;1883年德國動物學家魏斯曼稱之為種質。實際上魏斯曼所說的種質已經不再是單純的臆測了,他已經指明生殖細胞的染色體便是種質,並且明確地區分種質和體質,認為種質可以影響體質,而體質不能影響種質,在理論上為遺傳學的發展開闢了道路
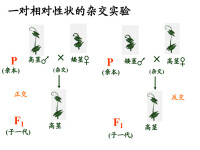
孟德爾三大定律
從1910年到現在遺傳學的發展大致可以分為三個時期:細胞遺傳學時期、微生物遺傳學時期和分子遺傳學時期。
遺傳學
這一時期通過對遺傳學規律和染色體行為的研究確立了遺傳的染色體學說。摩爾根在1926年發表的《基因論》和英國細胞遺傳學家達林頓在1932年發表的《細胞學的最新成就》兩書是這一時期的代表性著作。這一時期中雖然在1927年由美國遺傳學家米勒和1928年斯塔德勒分別在動植物中發現了 X射線的誘變作用,可是對於基因突變機制的研究並沒有進展。基因作用機制研究的重要成果則幾乎只限於動植物色素的遺傳研究方面。

遺傳學
在這一時期中,採用微生物作為材料研究基因的原初作用、精細結構、化學本質、突變機制以及細菌的基因重組、基因調控等,取得了已往在高等動植物研究中難以取得的成果,從而豐富了遺傳學的基礎理論。1900~1910年人們只認識到孟德爾定律廣泛適用於高等動植物,微生物遺傳學時期的工作成就則使人們認識到遺傳學的基本規律適用於包括人和噬菌體在內的一切生物。
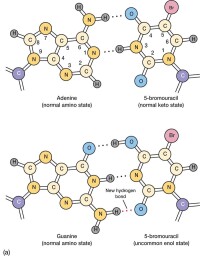
遺傳學
分子遺傳學是在微生物遺傳學和生物化學的基礎上發展起來的。分子遺傳學的基礎研究工作都以微生物、特別是以大腸桿菌和它的噬菌體作為研究材料完成的;它的一些重要概念如基因和蛋白質的線性對應關係、基因調控等也都來自微生物遺傳學的研究。分子遺傳學在原核生物領域取得上述許多成就后,才逐漸在真核生物方面開展起來。
正像細胞遺傳學研究推動了群體遺傳學和進化遺傳學的發展一樣,分子遺傳學也推動了其他遺傳學分支學科的發展。遺傳工程是在細菌質粒和噬苗體以及限制性內切酶研究的基礎上發展起來的,它不但可以應用於工、農、醫各個方面,而且還進一步推進分子遺傳學和其他遺傳學分支學科的研究。
免疫學在醫學上極為重要,已有相當長的歷史。按照一個基因一種酶假設,一個生物為什麼能產生無數種類的免疫球蛋白,這本身就是一個分子遺傳學問題。自從澳大利亞免疫學家伯內特在 1959年提出了克隆選擇學說以後,免疫機制便吸引了許多遺傳學家的注意。目前免疫遺傳學既是遺傳學中比較活躍的領域之一,也是分子遺傳學的活躍領域之一。
在分子遺傳學時代另外兩個迅速發展的遺傳學分支是人類遺傳學和體細胞遺傳學。自從採用了微生物遺傳學研究的手段后,遺傳學研究可以不通過生殖細胞而通過離體培養的體細胞進行,人類遺傳學的研究才得以迅速發展。不論研究的對象是什麼,凡是採用組織培養之類方法進行的遺傳學研究都屬於體細胞遺傳學。人類遺傳學的研究一方面廣泛採用體細胞遺傳學方法,另一方面也愈來愈多地應用分子遺傳學方法,例如採用遺傳工程的方法來建立人的基因文庫並從中分離特定基因進行研究等。
遺傳學
一個受精卵通過有絲分裂而產生無數具有相同遺傳組成的子細胞,它們怎樣分化成為不同的組織是一個遺傳學課題,有關這方面的研究屬於發生遺傳學。由一個受精卵產生的免疫恬性細胞能夠分別產生各種不同的抗體球蛋白,這也是遺傳學的一個課題,它的研究屬於免疫遺傳學。
DNA通過信使RNA作為中間載體編碼蛋白質。
血紅蛋白能夠在哺乳動物血液中運輸氧氣。圖中顯示了血紅蛋白在攜氧和脫氧狀態之間的結構變化。
單個氨基酸突變導致血紅蛋白形成纖維。基因通常是通過生成所編碼的蛋白質(執行細胞中大多數功能的複雜的生物大分子)來表現它們的功能性影響。蛋白質是由氨基酸所組成的線性鏈,而基因的DNA序列(通過RNA作為信息的中間載體)被用於產生特定的蛋白質的氨基酸序列。這一過程的第一步是由基因的DNA序列來生成一個序列互補的RNA分子,即基因的轉錄。
通過轉錄產生的RNA分子(信使RNA)被用於生產相應的氨基酸序列,這一轉換過程被稱為翻譯。核酸序列中的每一組三個核苷酸組成一個密碼子,可以被翻譯為20種出現於蛋白質中的氨基酸中的一個,這種對應性被稱為遺傳密碼。這種信息的傳遞是單一方向性的,即信息只能從核苷酸序列傳遞到氨基酸序列,而不能從氨基酸序列傳遞迴核苷酸序列,這一現象被弗朗西斯·克里克稱為分子生物學中心法則。
特定的氨基酸序列決定了對應蛋白質的獨特的三維結構,而蛋白質結構則與它們的功能緊密相連。一些蛋白質是簡單的結構分子,如形成纖維的膠原蛋白。蛋白質可以與其他蛋白質或小分子結合;例如,作為酶的蛋白質通過與底物分子結合來執行催化其化學反應的功能。蛋白質結構是動態的;例如,血紅蛋白在哺乳動物血液中捕捉、運輸和釋放氧氣分子的過程中能夠發生微小的結構變化。
也有一些基因被轉錄為RNA分子后卻不被翻譯成蛋白質,這些RNA分子就被稱為非編碼RNA。在一些例子中,這些非編碼RNA分子(如核糖體RNA和轉運RNA)摺疊形成結構並參與部分關鍵性細胞功能。還有的RNA(如microRNA)還能夠通過與其他RNA分子雜交結合而發揮調控作用。基因序列上的單個核苷酸變化(密碼子改變)可能會導致所編碼蛋白質的氨基酸序列相應改變。由於蛋白質結構是由其氨基酸序列所決定的,一個氨基酸的變化就有可能通過使結構失去穩定性或改變蛋白質表面而影響與該蛋白質其他蛋白質和分子的相互作用,而引起蛋白質性質發生劇烈的改變。例如,鐮刀型細胞貧血症是一種人類遺傳性疾病,是由編碼血紅蛋白中的β-球蛋白亞基的基因中的一個核苷酸突變所引起的,這一突變導致一個氨基酸發生改變從而改變了血紅蛋白的物理性質;在這一疾病中,突變的血紅蛋白互相結合在一起,堆積而形成纖維,從而扭曲了攜帶血紅蛋白的紅血球的形狀。這些扭曲的鐮刀狀細胞無法在血管中通暢地流動,容易堆積而阻塞血管或者被降解,從而引起貧血疾病。
在人類遺傳疾病苯丙酮尿症中,環境因素也具有重大的影響。導致苯丙酮尿症的突變破壞了機體降解苯丙氨酸的能力,導致具有毒性的中間產物分子在體內堆積,從而引起嚴重的進行性智能發育不全和癲癇。帶有苯丙酮尿症突變的病人需要遵守嚴格的飲食,以避免攝入含苯丙氨酸的食品,才能保持正常的和健康的生活。暹羅貓具有溫度敏感型突變,從而引起毛色的不同。雖然基因含有生物體所需功能的所有信息,環境依然在確定生物體最終的表現型中發揮著重要作用;這種兩面性被稱為“先天與後天”,也可以說,一個生物體的表現型依賴於遺傳與環境的相互作用。這種相互作用的一個例子就是溫度敏感型突變:蛋白質序列中的單個氨基酸突變通常不會改變該蛋白質的行為和與其他分子的相互作用關係,但卻能夠使該蛋白質結構變得不穩定。在一個高溫環境中,分子的運動加快,分子間的碰撞也加強,這就使得這一蛋白質的結構被破壞從而無法發揮它的功能;而在一個低溫環境中,蛋白質結構卻可以保持穩定並能夠發揮正常的功能。這類突變所引起的改變在暹羅貓毛色的變化中可以被觀察到。這種貓體內一種負責生產色素的酶含有一個突變,這個突變能夠導致這種酶在高溫時變得不穩定並失去其功能。因此,在貓皮膚溫度較低處(如四肢、尾部、面部等)的毛色為深色,而較高處為淺色。
多細胞生物中的基因表達的差異性非常明顯:雖然各類細胞都含有相同的基因組,卻由於不同的基因表達而具有不同的結構和行為。多細胞生物中的所有細胞都來源於一個單一細胞,通過響應外部或細胞之間的信號而不斷分化並逐漸建立不同的基因表達規律來產生不同的行為。因為沒有一個單一基因能夠負責多細胞生物中的各個組織的發育,因此這些規律應來自於許多細胞之間的複雜的相互作用。這些過程都要通過基因調控來完成。轉錄因子與DNA結合,影響了所結合基因的轉錄。一個生物體的基因組含有數千個基因,但並不是所有的基因都需要保持激活狀態。基因的表達表現為被轉錄為mRNA,然後再被翻譯成蛋白質;而細胞中存在許多方式可以來控制基因的表達,以便使蛋白質的產生符合細胞的需求。而控制基因表達“開關”的主要調控因子之一就是轉錄因子;它們是一類結合在基因的起始位點上的調控蛋白,可以激活或抑制基因的轉錄。例如,在大腸桿菌細菌基因組內存在著一系列合成色氨酸所需的基因。然而,當細菌細胞可以從環境中獲得色氨酸時,這些基因就不被細胞所需要。色氨酸的存在直接影響了這些基因的活性,這是因為色氨酸分子會與色氨酸操縱子(一種轉錄因子)結合,引起操縱子結構變化,使得操縱子能夠結合到合成色氨酸所需基因上。色氨酸操縱子阻斷了這些基因的轉錄和表達,因而對色氨酸的合成進程產生了負反饋調控作用。
真核生物體內的染色質中存在著能影響基因轉錄的結構特點,常常表現為DNA和染色質的修飾形式(如DNA的甲基化),而且能夠穩定遺傳給子細胞。這些特點是“附加性”的,因為它們存在於DNA序列的“頂端”並且可以從一個細胞遺傳給它的下一代。由於這些附加性特點,在相同培養基中生長的不同的細胞類型依然保持其不同的特性。雖然附加性特點在整個發育過程中通常是動態的,但是有一些,例如副突變(paramutation)現象可以被多代遺傳,也是DNA是遺傳的分子基礎這一通用法則的極少數例外。
遺傳學與生物化學的關係最為密切,和其他許多生物學分支學科之間也有密切關係。例如發生遺傳學和發育生物學之間的關係;行為遺傳學同行為生物學之間的關係;生態遺傳學同生態學之間的關係等。此外,遺傳學和分類學之間也有著密切的關係,這不僅因為在分類學中應用了DNA鹼基成分和染色體等作為指標,而且還因為物種的實質也必須從遺傳學的角度去認識。

遺傳學
美國在20年代中應用雜種優勢這一遺傳學原理於玉米育種而取得顯著的增產效果;中國在70年代把此原理成功地推廣應用於水稻生產。多倍體的生長優勢同樣在中國得到了應用,小黑麥異源多倍體的培育成功便是一例。人工誘變也是廣泛應用的育種方法之一。數量遺傳學和生物統計遺傳學的研究結果,被應用到動、植物選種工作中而使育種效率得以提高。這些主要是細胞遺傳學時期研究成果的應用。
40年代初,抗菌素工業的興起推動了微生物遺傳學的發展,微生物遺傳學的發展又推動了抗菌素工業以及其他新興的發酵工業的進步。隨著微生物遺傳學研究的深入,基因調控作用的原理被成功地應用到氨基酸等發酵工業中。此外雜交轉導、轉化等技術的採用也增加了育種的手段。
70年代體細胞遺傳學的發展進一步增加了育種的手段,包括所謂單倍體育種以及通過體細胞誘變和細胞融合的育種等。這些手段的應用將有可能大大地加速育種工作的進程。
遺傳學研究同人類本身密切相關。由於人類遺傳學研究的開展,特別是應用體細胞遺傳學和生化遺傳學方法所取得的進展,對於遺傳性疾病的種類和原因已經有很多了解;產前診斷和嬰兒的遺傳性疾病診斷已經逐漸推廣;對於某些遺傳性疾病的藥物治療也在研究中。免疫遺傳學是組織移植和輸血等醫學實踐的理論基礎;藥物遺傳學和藥物學有密切的關係;毒理遺傳學關係到藥物的安全使用和環境保護。用遺傳工程技術對遺傳性疾病進行基因治療也正在進行探索。人類遺傳學研究也是優生學的基礎。
遺傳學研究為致癌物質的檢測提供了一系列的方法。雖然目前治療癌症還沒有十分有效的方法,但在環境污染日益嚴重的今天能夠有效地檢測環境中的致癌物質,便是一個重大的進展。癌症患病的傾向性是遺傳的,癌症的起因又同DNA損傷修復有關,近年來癌基因的發現進一步說明癌症和遺傳的密切關係,所以從長遠觀點來看,遺傳學研究必將為全面控制癌症作出貢獻。
許多遺傳學分支的研究都採用了分子遺傳學手段,特別是重組DHA技術。即使是有關群體的遺傳學研究也受分子遺傳學的影響,進化遺傳學研究中的分子進化領域便是一個例子。
近幾年來,人類基因組研究的進展日新月異,而分子生物學技術也不斷完善,隨著基因組研究向各學科的不斷滲透,這些學科的進展達到了前所未有的高度。在法醫學上,STR位點和單核苷酸(SNP)位點檢測分別是第二代、第三代DNA分析技術的核心,是繼RFLPs(限制性片段長度多態性)VNTRs(可變數量串聯重複序列多態性)研究而發展起來的檢測技術。作為最前沿的刑事生物技術,DNA分析為法醫物證檢驗提供了科學、可靠和快捷的手段,使物證鑒定從個體排除過渡到了可以作同一認定的水平,DNA檢驗能直接認定犯罪、為兇殺案、強姦殺人案、碎屍案、強姦致孕案等重大疑難案件的偵破提供準確可靠的依據。隨著DNA技術的發展和應用,DNA標誌系統的檢測將成為破案的重要手段和途徑。此方法作為親子鑒定已經是非常成熟的,也是國際上公認的最好的一種方法。
1991-1997年,中國曾邦哲[傑](Zeng BJ.)發表《結構論-泛進化理論》系列論文,闡述系統醫藥學(systems medicine)、系統生物工程(system biological engineering)與系統遺傳學(system genetics)的概念,提出經典、分子與系統遺傳學發展觀,以及於2003年、2008年於國際遺傳學大會,採用結構(structure)、系統(system)、圖式(pattern)遺傳學的辭彙來描述系統科學方法、計算機技術研究生物系統遺傳結構、生物系統形態圖式之間的“基因型-表達型”複雜系統研究領域,以細胞信號傳導、基因調控網路為核心研究細胞進化、細胞發育、細胞病理、細胞藥理的細胞非線性系統動力學。
2003年挪威科學家稱之為整合遺傳學(integrative genetics)並建立了研究中心,2005年,國際上Cambien F.和 Laurence T.發表動脈硬化研究的系統遺傳學觀,Morahan G., Williams RW.等2007年(Bock G., Goode J. Eds.)論述系統遺傳學將成為下一代遺傳學。2005-2008年,國際系統遺傳學飛速發展,歐美建立了許多系統遺傳學研究中心和實驗室。2008年在美國召開了整合與系統遺傳學國際會議,2009年荷蘭舉辦了系統遺傳學國際會議,2008年美國國立衛生研究院(NIH)設立了腫瘤的系統遺傳學研究專項基金。系統遺傳學,採用計算機建模、系統數學方程、納米高通量生物技術、微流控晶元實驗等方法,研究基因組的結構邏輯、基因組精細結構進化、基因組穩定性、生物形態圖式發生的細胞發生非線性系統動力學。
模式生物
黑腹果蠅(Drosophila melanogaster)是一種流行於遺傳學研究中的模式生物。
一開始遺傳學家們的研究對象很廣泛,但逐漸地集中到一些特定物種(模式生物)的遺傳學上。這是由於新的研究者更趨向於選擇一些已經獲得廣泛研究的生物體作為研究目標,使得模式生物成為多數遺傳學研究的基礎。模式生物的遺傳學研究包括基因調控以及發育和癌症相關基因的研究。
模式生物具有傳代時間短、易於基因操縱等優點,使得它們成為流行的遺傳學研究工具。目前廣泛使用的模式生物包括:大腸桿菌(Escherichia coli)、釀酒酵母(Saccharomyces cerevisiae)、擬南芥(Arabidopsis thaliana)、線蟲(Caenorhabditis elegans)、果蠅(Drosophila melanogaster)以及小鼠(Mus musculus)。
遺傳學研究
醫學遺傳學的目的是了解基因變異與人類健康和疾病的關係。當尋找一個可能與某種疾病相關的未知基因時,研究者通常會用遺傳連鎖和遺傳系譜來定位基因組上與該疾病相關的區域。在群體水平上,研究者會採用孟德爾隨機法來尋找基因組上與該疾病相關的區域,這一方法也特別適用於不能被單個基因所定義的多基因性狀。一旦候選基因被發現,就需要對模式生物中的對應基因(直系同源基因)進行更多的研究。對於遺傳疾病的研究,越來越多發展起來的研究基因型的技術也被引入到藥物遺傳學中,來研究基因型如何影響藥物反應。
癌症雖然不是傳統意義上的遺傳病,但被認為是一種遺傳性疾病。癌症在機體內的產生過程是一個綜合性事件。機體內的細胞在分裂過程中有一定幾率會發生突變。這些突變雖然不會遺傳給下一代,但會影響細胞的行為,在一些情況下會導致細胞更頻繁地分裂。有許多生物學機制能夠阻止這種情況的發生:信號被傳遞給這些不正常分裂的細胞並引發其死亡;但有時更多的突變使得細胞忽略這些信號。這時機體內的自然選擇和逐漸積累起來的突變使得這些細胞開始無限制生長,從而成為癌症性腫瘤(惡性腫瘤),並侵染機體的各個器官。
研究技術
瓊脂平板上的大腸桿菌菌落,細胞克隆的一個例子,常用於分子克隆。
可以在實驗室中對DNA進行操縱。限制性內切酶是一種常用的剪切特異性序列的酶,用於製造預定的DNA片斷。然後利用DNA連接酶將這些片斷重新連接,通過將不同來源地DNA片斷連接到一起,就可以獲得重組DNA。重組DNA技術通常被用於在質粒(一種短的環形DNA片斷,含有少量基因)中,這常常與轉基因生物的製造有關。將質粒轉入細菌中,再在瓊脂平板培養基上生長這些細菌(來分離菌落克隆),然後研究者們就可以用克隆菌落來擴增插入的質粒DNA片斷(這一過程被稱為分子克隆)。
DNA還能夠通過一個被稱為聚合酶鏈鎖反應(又被稱為PCR)的技術來進行擴增。利用特定的短的DNA序列,PCR技術可以分離和擴增DNA上的靶區域。因為只需要極少量的DNA就可以進行擴增,該技術也常常被用於DNA檢測(檢測特定DNA序列的存在與否)。
測序與基因組
DNA測序技術是遺傳學研究中發展起來的一個最基本的技術,它使得研究者可以確定DNA片段的核苷酸序列。由弗雷德里克·桑格和他的同事於1977年發展出來的鏈終止測序法現在已經是DNA測序的常規手段。在這一技術的幫助下,研究者們能夠對與人類疾病相關的DNA序列進行研究。
由於測序已經變得相對廉價,而且在計算機技術的輔助下,可以將大量不同片斷的序列信息連接起來(這一過程被稱為“基因組組裝”),因此許多生物(包括人類)的基因組測序已經完成。這些技術也被用在測定人類基因組序列,使得人類基因組計劃得以在2003年完成。隨著新的高通量測序技術的發展,DNA測序的費用被大大降低,許多研究者希望能夠將測定一個人的基因組信息的價格降到一千美元以內,從而使大眾測序成為可能。
一個或多個基因異常,特別是隱性基因,是相當普遍的。每個人都攜帶有6~8個異常隱性基因。然而,這些基因並不引起細胞功能異常,除非有兩個相似的隱性基因存在。一般人群中,具有兩個相似隱性基因的個體機率非常小,但是在近親婚配的孩子中,這種機率較高。在近親婚配的群體中,比如基督教的阿曼門諾派,機率也較高。
一個人的基因組成稱之為基因型。這些基因在人體存在的反映,即基因型的表達稱之為表現型。
所有遺傳特徵(性狀)為基因所編碼。有一些特徵,比如頭髮顏色、人與人之間的差異等,不能算是異常,然而,由異常基因表達產生的異常特性可能導致遺傳疾病。
單基因異常
單個基因異常的影響取決於這個基因是顯性還是隱性基因,和這個基因是否位於X染色體上(X連鎖)。因為一個基因指導一種特定蛋白質的合成,因此異常基因就可能產生異常蛋白質或導致某種蛋白質含量異常,進而引起細胞功能的異常,最終導致機體形態或功能異常。
染色體遺傳
位於常染色體上的異常顯性基因產生的性狀,可以導致畸形、疾病或有發展成某一疾病的傾向。
下面一些原則通常適用於顯性基因決定的性狀:
有這種性狀的人,至少父母有一方具有同樣性狀,除非這種性狀是由新的基因突變引起。
異常遺傳性狀常常是由新的遺傳突變所致,而不是由父母遺傳。
當父母一方具有異常性狀,而另一方沒有時,他們的每一個孩子有50%的機率遺傳這種異常,有50%的機率沒有遺傳這種性狀。如果父母之一具有兩個這種異常基因(雖然很罕見),他們所有子女都將有異常性狀。
不具異常性狀的個體,即使他的兄弟姊妹有異常性狀,因為他不攜帶這種基因,他的子女也不會遺傳異常性狀。
男性和女性受影響的可能性相同。
常常在每一代中都出現異常。
下列一些原則通常適用於由隱性基因決定的性狀:
實際上,具有這種性狀的人,其父母雙方都可能攜帶有這種隱性基因,但父母雙方均不表現出這種性狀。
突變很少引起該性狀的表達。
父母之一表現出性狀,另一方攜帶隱性基因,沒有表現性狀,他們的子女中約有一半有可能表現出異常性狀;而另一半將是隱性基因的攜帶者。假如另一方沒有攜帶異常基因,他們的子女將無一人表現出異常性狀,但他們所有的子女都將是攜帶者,並且可將這個異常基因遺傳給他們子女的後代。
一個沒有異常性狀的人,如果兄弟姊妹有異常性狀,他很可能攜帶有一個這種異常基因。
男性和女性受影響的可能性相同。
通常不是在每一代都有異常表現,除非父母雙方均表現為異常。
引起嚴重疾病的顯性基因是罕見的。因為這類基因趨於消失,帶有引起嚴重疾病顯性基因的人常常因為疾病太嚴重而沒有生育能力。然而,有少數例外,如亨廷頓舞蹈病(見第67節),可引起嚴重的腦功能衰退,癥狀常開始於35歲之後,在癥狀表現之前,患者可能已經有了孩子。
隱性基因僅僅是在該個體具有兩個同樣的這種隱性基因時才有可能表達。僅有一個隱性基因的個體不表現出性狀,但他是這種隱性基因的攜帶者,能夠將此基因遺傳給他的孩子。
X連鎖基因
因為男性的Y染色體上的基因數目很少,在單個X染色體上的基因(X連鎖基因)都是不成對的。因此,無論是顯性基因或是隱性基因都能夠表達。但是,女性有兩條X染色體,適用於常染色體上基因的那些原則,同樣適用於女性X連鎖基因。除非成對的兩個基因都是隱性的,通常只有顯性基因才能表達。
假如一個異常X連鎖基因是顯性,受影響的男性會將異常基因遺傳給他所有的女兒,但不會遺傳給兒子。受影響的男性的兒子接受他的Y染色體,而Y染色體不攜帶異常基因。只帶有一個異常基因的女性,可以把異常基因遺傳給她的一半子女,兒子或女兒。
假如異常X連鎖基因是隱性基因,幾乎出現異常性狀的均為男性。異常的男性只把異常基因傳給女兒,她們都是攜帶者。攜帶者母親並不表現異常,但可將異常基因遺傳給一半兒子,這些兒子常常表現出異常性狀。她們的女兒沒有異常性狀,但女兒中有一半是攜帶者。
紅綠色盲,由X連鎖隱性基因引起,男性發生率大約為10%,而女性則不常見。在男性,引起色盲的基因來自母親,母親是色盲患者或者是具有正常視力的色盲基因攜帶者。色盲基因不來自父親,因為父親提供Y染色體。色盲父親的女兒極少是色盲,但常常是色盲基因的攜帶者。
等顯性遺傳
等顯性遺傳,兩個基因均表達。例如鐮狀細胞貧血;假如一個人有一個正常基因,同時又有一個異常基因,就可能產生正常和異常兩種血紅蛋白。
線粒體基因
每一個細胞內都有線粒體,這是一種提供細胞能源的微小結構。每個線粒體含有一個環形染色體。有幾種罕見的疾病為線粒體內染色體攜帶的異常基因所引起。
當卵細胞受精時,只有卵細胞的線粒體成為發育中胎兒的一部分;精子的所有線粒體均被拋棄。因此,異常線粒體基因所致的疾病是通過母親遺傳的。而具有異常線粒體基因的父親,不會遺傳這類疾病給他的孩子們。
癌細胞可能含有致癌基因,致癌基因是引起癌症的基因(也稱腫瘤基因)(見第162節)。有時癌基因是出生前負責生長發育的基因的異常翻譯,正常情況下,出生后它們就永遠失活。如果這些致癌基因以後被再激活,可能導致癌症發生。它們是如何被再激活的尚不知道。
基本信息
- 中文名
- 遺傳學
- 外文名
- Genetics
- 研究對象
- 父母子女或一個家族
- 研究範圍
- 遺傳與變異的科學
- 重要物質
- DNA
- 單位
- 細胞為單位