著絲粒
染色體中連接兩個染色單體並將染色單體分為短臂(p) 和長臂(q)的結構
著絲粒是染色體中連接兩個染色單體並將染色單體分為短臂(p) 和長臂(q)的結構。由於著絲粒區染色質較細、內縊,所以也稱主縊痕(primary constriction),此處DNA 高度重複,為鹼性染料所深染。
物理功能上,著絲粒為動粒組裝提供了位點。動粒是實際上負責染色體分離的一種高度複雜的蛋白質結構。當所有染色體都與紡錘體以合適的方式結合之後,結合微管蛋白並向細胞發出信號,以保證細胞的正確分裂。
廣義上,生物中的著絲粒可以分為兩種。“點著絲粒”與特定蛋白質結合,並高效的特異性識別DNA序列。在特定物種中,任何具有點著絲粒序列的DNA都通常形成著絲粒結構。點著絲粒在釀酒酵母(Saccharomyces cerevisiae)中表徵最為明顯。“區域著絲粒”則用來描述其餘的大多數著絲粒,通常在DNA上具有偏好性的區域形成。形成區域著絲粒的信號似乎是表觀遺傳的。大多數生物(從裂殖酵母到人類)都具有區域著絲粒。
具體在有絲分裂過程中,著絲粒代表一對姐妹染色單體之間接觸最近的區域。當細胞進入分裂期后,一對姐妹染色單體沿長度方向通過黏連蛋白複合體相互連接。這種複合物被認為是在前期由染色體臂釋放出的,因此當染色體排列在有絲分裂中紡錘體中間板上時,它們最後相連的位置在著絲粒以及附近的染色質中。
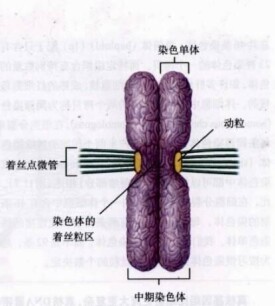
著絲粒位於異染色質區內,這裡富集了衛星DNA,也就是短的DNA串聯重複序列。此外,在縊痕區內有一個直徑或長度為400 nm左右的很緻密的顆粒狀結構,這稱為動粒(kinetochore)的結構直接與牽動染色體向兩極移動的纖絲蛋白相連結。
在染色體上,著絲粒有多種可能的存在位置。一般來講,主要的位置有中間著絲,亞中間著絲,近端著絲,端著絲。
著絲粒位於染色體的中心,染色體兩臂幾乎等長,形成"X"形狀。在人類染色體組中,第1,3,16,19,20號染色體是中間著絲的。有些情況下,中間著絲通過兩個近端著絲染色體的融合形成。
如果與中間著絲類似,而染色體兩臂不等長,則為亞中間著絲。Y染色體是亞中間著絲的。
如果染色體短臂存在,但由於過短而難以觀察,則成為近端著絲。人類染色體組中13,14,15,21,22號為近端著絲。在這種著絲情況下,染色體短臂(p)內仍然包含遺傳物質,如核仁組織區域。
端著絲情況下著絲粒位於染色體的末端。兩條染色體的端粒從末端延伸並結合。此時染色體只有一條臂。典型例子在家鼠的染色體組中出現。
除這幾種常見著絲方式以外,也存在其他組織方式。如全著絲,整條染色體都作為著絲粒區域,最知名的例子是秀麗隱桿線蟲(Caenorhabditis elegans)。無中心,此時染色體缺少著絲粒。
著絲粒區域一般處於異染色質狀態,這對於其對黏連蛋白複合體的招募十分重要。在這種染色質中,一般的組蛋白H3被另外的中心粒特異性蛋白(人類中為CENP-A)代替。 CENP-A被認為對動粒在著絲粒上的組裝起重要作用。研究發現CENP-C幾乎專一地定位於結合CENP-A的染色質區域。在著絲粒區域中,對於人類,其組蛋白的主要修飾為H4K20me3和H3K9me3。
在裂殖酵母中,著絲粒異染色質的形成與RNAi有關。在線蟲類,一些植物,以及半翅類,鞘翅類昆蟲中,染色體主要以全著絲的形式存在,表明其不存在優先的微管蛋白結合位點。
染色體著絲粒(centromere)的主要作用是使複製的染色體在有絲分裂和減數分裂中可均等地分配到子細胞中。在很多高等真核生物中,著絲粒看起來像是在染色體一個點上的濃縮區域,這個區域包含著絲點 (希臘語 kínesis 運動; chóros 部位),又稱主縊痕。此是細胞分裂時紡錘絲附著之處。在大部分真核生物中每個紡錘絲附著在不同的著絲粒上。如啤酒酵母(Saccharomyces cerevisiae)附著在每個著絲粒上僅一條紡錘絲。廣義上說著絲粒也常指著絲點﹐然而狹義上的著絲點是將染色體和紡錘絲微管相結合的蛋白質複合體。
若著絲粒丟失了,那麼染色體就失去了附著到紡錘絲上的能力,細胞分裂時染色體就會隨機地進入子細胞。然而有著絲粒的染色體也會出現這種異常分配,那就是複製后的兩個染色體拷貝並不總是正確地分離進入子細胞。在此過程中發生錯誤的概率通常是很低的。若發生錯誤會引起染色體數目的改變。如在酵母中分配發生錯誤的概率低於十萬分之一。
由於著絲粒DNA序列並不是動物中著絲粒的關鍵決定因素,因此表觀遺傳被認為在著絲粒的特化中起關鍵作用。染色體分裂形成的子染色體會與形成其的染色體在相同的位置形成著絲粒,而與序列無關。目前假設H3組蛋白變體CENP-A是著絲粒的主要表觀遺傳標誌。
著絲點是高中生物學教科書常用的染色體基本結構名稱。本套教科書在第1冊有絲分裂和減數分裂有關細胞分裂中均用“著絲點”,而在第2冊染色體組型分析中對染色體分類卻用“著絲粒”。許多學生疑問“著絲點和著絲粒有什麼區別?是不是同一結構?”
經查,著絲點為Kinetochore,著絲粒為Centromere,在許多文獻資料中使用不一。例如,在《細胞生物學》(1987年,高等教育出版社)中二者均有使用,劉祖洞和江紹慧的《遺傳學》(1987年,高等教育出版社)中只用“著絲粒”,中央農業廣播電視學校教材《植物及植物生理》(修訂執筆人孟繁靜等,1989年,農業出版社)中只用“著絲點”。近來在電鏡下觀察發現的資料表明,著絲粒(染色體的主縊痕primary constriction)為染色質的結構,將染色體分成二臂,在細胞分裂前期和中期,把兩個姐妹染色單體連在一起,到後期兩個染色單體的著絲粒分開。著絲粒兩側各有一個由蛋白質構成的3層盤狀特化結構,為非染色體性質物質的附加物,稱為著絲點,在染色質(染色體)被鹼性染料染色時,著絲點部分染色很淺或根本不染色,由於著絲點部位幾乎把著絲粒覆蓋,所以,染色后觀察染色體的外形,在著絲點部位幾乎看不到著色。著絲點與染色體的移動有關,在細胞分裂(包括有絲分裂和減數分裂)的前、中、後期,紡錘體的紡錘絲(或星射線)微管就附著在著絲點上,並牽引染色體移動,意即紡錘體的紡錘絲(或星射絲)直接附著在著絲點上而不是附著在染色體著絲粒上,沒有著絲點,染色體不能由紡錘絲牽引移動。因此,著絲點和著絲粒並非同一結構,它們的功能也不同,但它們的位置關係是固定的,有時用著絲點或著絲粒泛指它們所在的染色體主縊痕位置是可以理解的。
根據人民衛生出版社出版的七年制臨床醫學教材《細胞生物學》,在兩條姐妹染色單體相連處,有一個向內凹陷的縊痕,稱為主縊痕(primary constriction),光鏡下相對不著色。著絲粒處於主縊痕的內部,是主縊痕的染色質部位。電鏡下可見主縊痕兩側有一三層結構的特化部位,稱為動粒(kinetochore),即著絲點。目前認為著絲粒是染色單體中一段高度重複的DNA序列,該序列不與組蛋白結合。動粒是著絲粒結合蛋白在有絲分裂染色體著絲粒部位形成的一種圓盤狀的結構,微管與之連接,與染色體分離密切相關,每一個中期染色體有兩個動粒,位於著絲粒的兩側。
動粒又叫著絲點,是附著於著絲粒上的一種細胞結構。動粒可分為內板、中間間隙、外板和纖維冠4個部分。在細胞分裂過程中,微管與動粒相連,牽引染色體在分裂中期進行染色體列隊,在分裂後期,牽引分開的染色體分別向細胞的兩極運動。
目前正在研究著絲粒結合蛋白以及其它的一些因素。一個主要的問題是解決紡錘絲附著到著絲粒的具體機制。
基本信息
- 中文名
- 著絲粒
- 外文名
- centromere
- 活動
- 有絲分裂(mitosis)和減數分裂(
- 組成
- 高度重複DNA序列的異染色質
- 細胞
- 真核生物細胞