細胞壁
生物術語
細胞壁是一個生物術語,拼音為xì bāo bì。細胞壁屬於細胞結構,是位於細胞膜外的一層較厚、較堅韌並略具彈性的結構,其成分為黏質複合物,有的種類在壁外還具有由多糖類物質組成的莢膜,起保護作用。植物、真菌、藻類和原核生物都具有細胞壁,而動物細胞不具有細胞壁。細胞壁之厚薄常因組織、功能不同而異。
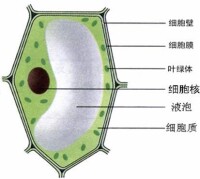
植物細胞
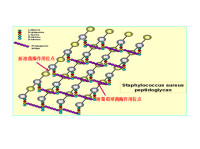
肽聚糖
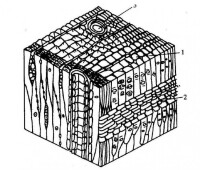
細胞壁
化石研究表明,大約在35億年前地球就已出現了原核細胞,大約在12~14億年前才出現真核細胞。關於真核細胞的起源,主要有兩種假說:一是“內共生假說”,認為真核細胞的各部分別起源於幾種共生的原核細胞,需氧細菌穿入異養厭氧的 原核生物變為線粒體,藍藻穿入變成葉綠體,螺旋體穿入變成鞭毛和纖毛等;一是“質膜內褶假說”,認為古原核生物隨著體積增大,質膜發生內褶而形成內質網等細胞器,以及核膜造成細胞核等;同時,DNA複製成許多拷貝,質膜圍繞著DNA發生內褶,最後形成有雙層膜的細胞器,如線粒體、葉綠體等。細胞的形狀和大小隨生物的種類而不同,即使同一生物不同部位細胞的形態也不相同。單獨存在的遊離細胞常呈球形或近似球形,但由於細胞表面張力 或原生質粘度及其不均勻性,細胞的外形有時也會發生變化。構成組織的細胞受相鄰細胞之間機械力和方向性的制約,往往呈現出不同的形態。一般說來,細胞形態與其生理功能密切相關。不同種類的細胞大小懸殊,細菌細胞一般直徑為0.5~1.5微米,長1~5微米,種子植物的幼嫩細胞直徑約為5~25微米,成熟細胞直徑均為15~65微米。高等動物的組織細胞通常比植物細胞小,但卵細胞一般都較大,目前已知最大的細胞是鴕鳥的卵細胞,直徑約5cm。有的神經細胞的突起可長達1米以上。最小的細胞為支原體細胞,直徑只有0.1微米。單細胞生物由一個細胞組成。多細胞生物體的細胞數目差別很大。人出生時約有 即百萬億個細胞;成人約有 ×16即1600萬億 個細胞,近百種類型。儘管生物細胞類型多樣,卻具有大致相同的基本結構,真核細胞的亞顯微結構可分為表面結構和內部結構兩大部分。表面結構包括細胞膜和膜外物質層,如植物細胞的細胞壁和某些動物細胞的細胞外被;內部結構包括細胞核和細胞質。細胞核由核膜、核質、核仁和染色質組成。細胞質中未分化的半透明膠態溶液稱為基質;具有特殊功能的各種微細結構稱為細胞器,如線粒體、質體、內質網、核糖體、高爾基體、溶酶體、中心體、微管、微絲,以及鞭毛和纖毛等;細胞質內常含有許多代謝產物形成的顆粒,如澱粉粒、糊粉粒、脂肪滴、糖原粒等,統稱后成質。細胞是生命活動的單位,一些生命活動的基本過程,如物質代謝、能量轉換、運動、發育、繁殖和遺傳等,都是以細胞為結構基礎來實現的。
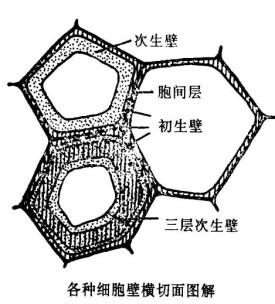
細胞壁分為3層,即胞間層(中層)、初生壁和次生壁。胞間層把相鄰細胞粘在一起形成組織。初生壁在胞間層兩側,所有植物細胞都有。次生壁在初生壁的裡面,又分為外(S1)、中(S2)、內(S3)3層,在內層裡面,有時還可出現一層。這樣的厚壁,水分和營養物就不能透過。有些植物的次生壁上具瘤層,還分化有特殊結構,如紋孔和瘤狀物等。紋孔是細胞間物質流通的區域,而瘤狀物則是次生壁裡層上的突起。
細胞壁的結構一般分下列三層
1.胞間層:胞間層是在細胞分裂產生新細胞時形成的,是相鄰兩個細胞間所共有的一層薄膜。它的主要成分是膠粒柔軟的果膠質。胞間層既將相鄰細胞粘連在一起,又可緩衝細胞間的擠壓,也不會阻礙細胞生長。
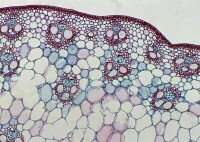
電鏡下的植物細胞壁
3.次生壁:細胞停止生長后,原生質體仍繼續分泌纖維素和其他物質,增添在初生壁內方,使細胞壁加厚,這部分加厚的細胞壁叫次生壁。次生壁添加在初生壁裡面,次生壁越厚,壁內的細胞腔就越小。次生壁只在植物體的部分細胞中有。厚壁的纖維細胞、石細胞、管胞和導管等有明顯增厚的次生壁。
細胞壁的主要組成成分是纖維素,它形成細胞壁的框架,內含其他物質。在電子顯微鏡下看到,這種框架由一層層纖維素微絲,簡稱微纖絲組成的,每一層微纖絲基本上是平行排列,每添加一層,微纖絲排列的方位就不同,因此層與層之間微纖絲的排列交錯成網。微纖絲之間的空間通常被其他物質填充。
細胞壁上有紋孔,是因為在細胞生長過程中,次生壁隨著細胞的生長而不斷 伸展,但壁的增厚是不均勻的,形成了許多壁薄的區域,叫做初生紋孔場;細胞產生次生壁時,增厚也不均勻,一般在初生紋孔場的部位不再加厚,細胞壁上就形成紋孔的結構。相鄰細胞壁上的紋孔常對應地形成紋孔對。紋孔有單紋孔和具緣紋孔兩種。通常有許多胞間連絲從紋孔通過,胞間連絲又跟細胞質中的內質網連接,從而溝通細胞間的物質交流,有利於水分的運輸。因此,細胞壁上的紋孔是細胞間聯繫的通道,使整個植物體在生命活動中能成為有機的統一體。
新細胞壁的形成是在細胞分裂末期的赤道面上,分裂的母細胞先形成成膜體。在染色體分向兩極時,高爾基器分離出的小泡與微管集合在赤道面上成為細胞板。新的多糖物質沉積在細胞板上就逐漸形成胞間層。其後細胞內合成一些纖維素組成微纖絲沉積在胞間層的兩側,就出現了初生壁。當細胞成熟停止生長以後,一層層新的纖維素和半纖維素以及木質素陸續添加在初生壁上,就建成了次生壁。初生壁每添加一層,微纖維排列的方向就可不同(縱向或橫向),形成了不規則的交錯網狀,稱為多網生長。這樣加厚的結果,使整個植物體的機械支持有了基礎。
細胞壁的胞間層基本上是由果膠質組成。
如果植物組織中的果膠質用果膠酶分解掉,細胞就會離散,這是因為初生壁是由水、半纖維素、果膠質、纖維素、蛋白質和脂類組成。胚芽鞘、莖、葉、毛等初生壁的各種成分的平均值見表。構成細胞壁的成分中,90%左右是多糖,10%左右是蛋白質、酶類以及脂肪酸。細胞壁中的多糖主要是纖維素、半纖維素和果膠類,它們是由葡萄糖、阿拉伯糖、半乳糖醛酸等聚合而成。次生細胞壁中還有大量木質素。
細菌細胞壁主要成分則是肽聚糖。
細胞壁的形成是多種細胞器配合作用的結果。新細胞壁的形成開始於細胞分裂的晚後期或早期。細胞分裂時,在兩組染色體之間,也就是在母細胞的赤道板(不是實際存在的)面上,有許多大小不一的分泌囊泡(secretoryvesicles)不規則地匯聚在一塊,這些小囊泡是由高爾基體和內質網分泌而形成的,其中富含組成細胞壁的各種糖類,它們藉助與細胞赤道板垂直方向上存在的微管的運動,逐漸整齊地排列成片,組成成膜體(phragmoplast)。成膜體中的囊泡膜相互融合與連接形成細胞的質膜,其中的內含物連成一體構成細胞板,這是雛形的中層結構。細胞板組成后,高爾基體小泡運輸造壁物質釋放到質膜外,以充實新形成的壁。當細胞板中逐漸有果膠質和少量纖維素分子不斷地填充和摻入時便構成了中層,在中層兩側陸續有纖維素和半纖維素等物質的沉積則形成了質地柔軟的初生壁,這時兩個子細胞便形成。此後,大多數細胞的初生壁內側又分層、定向地沉積著纖維素分子,它們經緯分明地交叉加固,這是增強植物體支持能力的重要基礎。纖維素分子的定向分層沉積與微管的活動有關,秋水仙素(colchicine)可阻止微管的形成,抑制纖維素分子的定向排列。微管的另一個重要作用是使新形成的細胞板上保留某些通道,即參與胞間連絲的形成,使原生質在兩個子細胞間能保持聯繫。
可見,細胞壁的形成是在生活細胞分裂、成長以至分化的過程中逐步完成的。在細胞分裂以及新細胞壁形成時,除了有高爾基體、內質網和微管參與外,還有生長素和多種酶類的作用,而所有的活動又要靠線粒體來提供能量,這正體現了細胞內各部位相互配合來共同完成生命活動的特徵。
細菌細胞壁、植物細胞壁、真菌細胞壁
細菌細胞壁主要成分是肽聚糖(peptidoglycan),又稱粘肽(mucopetide)。細胞壁的機械強度有賴於肽聚糖的存在。合成肽聚糖是原核生物特有的能力。肽聚糖是由n-乙醯葡萄糖胺和n-乙醯胞酸兩種氨基糖經β-1.4糖苷鍵連接間隔排列形成的多糖支架。在n-乙醯胞壁酸分子上連接四肽側鏈,肽鏈之間再由肽橋或肽鏈聯繫起來,組成一個機械性很強的網狀結構。各種細菌細胞壁的肽聚糖支架均相同,在四肽側鏈的組成及其連接方式隨菌種而異。
通常採用革蘭氏染色技術可以將細菌細胞壁區分為兩種類型,革蘭氏陽性(G+)和革蘭氏陰性(G-)。革蘭氏染色(Gram stain)是丹麥醫生革蘭(Hans Christian Gram)於1884年採用表2-3所列程序對細菌染色,結果因顯色不同可將細菌區分為兩類,分別稱為革蘭氏陽性和陰性。
革蘭氏陽性菌的四肽側鏈氨基酸由d-丙-d-谷-r-l-賴-d-丙組成。初合成的肽鏈末端多一個d-丙氨酸殘基。肽橋是一條5個甘氨酸的肽鏈,交聯時一端與側鏈第三位上賴氨酸連接,另一端在轉肽酶的作用下,使另一條五肽側鏈末端d-丙氨酸脫去,而與側鏈第四位d-丙氨酸連接。從x光檢查可見肽聚糖的多糖鏈是一條較硬而又呈螺旋狀捲曲的長桿,由於其呈螺旋狀,連接在其上的肽鏈才伸向四方,使交聯受到一定了限制,只有鄰近的肽鏈才可交聯。但葡萄球菌的肽橋較 長,有可塑性,使遠距離的肽鏈間也可交聯,交聯率達90%,形成堅固緻密的三維立體網狀結構。革蘭氏陰性大腸桿菌的四肽側鏈中第三位的氨基酸被二氨基庚二酸(dap)所取代,以肽鏈直接與相鄰四肽側鏈中的d-丙氨酸相連,且交聯率低,沒有五肽交聯橋,形成二維平面結構,所以其結構較革蘭氏陽性的葡萄球疏樺。
細菌細胞壁堅韌而富有彈性,保護細菌抵抗低滲環境,承受所在環境內的5~25個大氣的滲透壓,並使細菌在低滲的環境下細胞不易破裂;細胞壁對維持細菌的固有形態起重要作用;可允許水分及直徑小於1nm的可溶性小分子自由通過,與物質交換有關;細胞壁上帶有多種抗原決定簇,決定了細菌菌體的抗原性。
細菌細胞壁結構
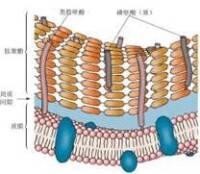
革蘭氏陽性菌細胞壁
革蘭氏陽性菌細胞壁 其細胞壁較厚,化學組成簡單,一般有90%肽聚糖和10%磷壁酸。細胞壁基本由厚厚一層肽聚糖構成,磷壁酸穿插其中。磷壁酸分兩類,一類與肽聚糖分子進行共價結合,稱壁磷壁酸,另一類是跨越肽聚糖層與細胞質膜相連,稱為膜磷壁酸或脂磷壁酸。細胞壁與細胞質膜之間的空間稱為周質間隙。
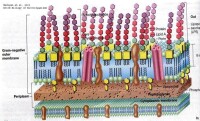
革蘭氏陰性菌細胞壁
酵母細胞壁 酵母細胞壁的厚度為0.1~0.3μm,重量占細胞乾重的18%~30%,主要由D-葡聚糖和D-甘露聚糖兩類多糖組成,含有少量的蛋白質、脂肪、礦物質。大約等量的葡聚糖和甘露聚糖占細胞壁乾重的 85%。當細胞衰老后,細胞壁重量會增加一倍。它雖然有一定韌性,但其堅韌性使酵母保持特殊的形狀。其化學成分較特殊,主要由酵母纖維素組成,它的結構類似三明治。外層為甘露聚糖,約佔細胞壁乾重的40%~45%。中間層是一層蛋白質分子,約佔細胞壁乾重的10%。其中有些是以與細胞壁相結合的酶的形式存在。內層為葡聚糖。
酵母葡聚糖是一種不溶性的有分支聚合物,主鏈以β-1,6糖苷鍵結合,支鏈以β-1,3糖苷鍵結合。作為細胞壁的內層物質,它維持細胞壁的強度,當細胞處於高滲的環境下而收縮時,它能維持細胞的彈性。
植物細胞的細胞壁主要成分是纖維素和果膠。植物細胞壁是植物細胞區別於動物細胞的主要特徵之一。由三部分組成:
1、胞間層
胞間層又稱中膠層。位於兩個相鄰細胞之間,為兩相鄰細胞所共有的一層膜,主要成分為果膠質。有助於將相鄰細胞粘連在一起,並可緩衝細胞間的擠壓。
2、初生壁
初生壁細胞分裂后,最初由原生質體分泌形成的細胞壁。存在於所有活的植物細胞。位於胞間層內側。通常較薄,約1~3微米厚。具有較大的可塑性,既可使細胞保持一定形狀,又能隨細胞生長而延展。主要成分為纖維素、半纖維素,並有結構蛋白存在。細胞在形成初生壁后,如果不再有新的壁層積累,初生壁便是他們的永久的細胞壁。如薄壁組織細胞。
3、次生壁
部分植物細胞在停止生長后,其初生壁內側繼續積累的細胞壁層。位於質膜和初生壁之間。主要成分為纖維素,並常有木質存在。通常較厚,約5~10微米,而且堅硬,使細胞壁具有很大的機械強度。大部分具次生壁的細胞在成熟時,原生質體死亡。纖維和石細胞是典型的具次生壁的細胞。在作植物原生質體培養時,常用含有果膠酶和纖維素酶的酶混合液處理植物組織,以破壞胞間層和去掉細胞的纖維素外壁,得到遊離的裸露原生質體。
真菌細胞壁中主要成分為幾丁質。
真菌細胞壁厚約 100~250nm, 它占細胞干物質的 30% 。細胞壁的主要成分為多糖,其次為蛋白質、類脂。在不同類群的真菌中,細胞壁多糖的類型不同。真菌細胞壁多糖主要有幾丁質 ( 甲殼質 ) 、纖維素、葡聚糖、甘露聚糖等,這些多糖都是單糖的聚合物,如幾丁質就是由 N- 乙醯葡萄糖胺分子,以 b -1 , 4 葡萄糖苷鍵連接而成的多聚糖。低等真菌的細胞壁成分以纖維素為主,酵母菌以葡聚糖為主,而高等真菌則以幾丁質為主。一種真菌的細胞壁組分並不是固定的,在其不同生長階段,細胞壁的成分有明顯不同。
酵母細胞壁:酵母細胞壁的厚度為0.1~0.3μm,重量占細胞乾重的18%~30%,主要由D-葡聚糖和D-甘露聚糖兩類多糖組成,含有少量的蛋白質、脂肪、礦物質。大約等量的葡聚糖和甘露聚糖占細胞壁乾重的85%。當細胞衰老后,細胞壁重量會增加一倍。它雖然有一定韌性,但其堅韌性使酵母保持特殊的形狀。其化學成分較特殊,主要由酵母纖維素組成,它的結構類似三明治。外層為甘露聚糖,約佔細胞壁乾重的40%~45%。中間層是一層蛋白質分子,約佔細胞壁乾重的10%。其中有些是以與細胞壁相結合的酶的形式存在。內層為葡聚糖。酵母葡聚糖是一種不溶性的有分支聚合物,主鏈以β-1,6糖苷鍵結合,支鏈以β-1,3糖苷鍵結合。作為細胞壁的內層物質,它維持細胞壁的強度,當細胞處於高滲的環境下而收縮時,它能維持細胞的彈性。
1、維持細胞形狀,控制細胞生長細胞壁增加了細胞的機械強度,並承受著內部原生質體由於液泡吸水而產生的膨壓,從而使細胞具有一定的形狀,這不僅有保護原生質體的作用,而且維持了器官與植株的固有形態。另外,壁控制著細胞的生長,因為細胞要擴大和伸長的前提是要使細胞壁鬆弛和不可逆伸展。
2、物質運輸與信息傳遞細胞壁允許離子、多糖等小分子和低分子量的蛋白質通過,而將大分子或微生物等阻於其外。因此,細胞壁參與了物質運輸、降低蒸騰作用、防止水分損失(次生壁、表面的蠟質等)、植物水勢調節等一系列生理活動。細胞壁上紋孔或胞間連絲的大小受細胞生理年齡和代謝活動強弱的影響,故細胞壁對細胞間物質的運輸具有調節作用。另外,細胞壁也是化學信號(激素、生長調節劑等)、物理信號(電波、壓力等)傳遞的介質與通路。
3、防禦與抗性細胞壁中一些寡糖片段能誘導植保素(phytoalexin)的形成,它們還對其它生理過程有調節作用,這種具有調節活性的寡糖片斷稱為寡糖素(oligosaccharin)。將一種庚葡萄糖苷寡糖素施加於大豆細胞時,會使負責合成抑制黴菌生長的抗菌素的基因活化而產生抗菌素。多種寡糖素的功能複雜多樣,如有的作為蛋白酶抑製劑誘導因子,在植物抵抗病蟲害中起作用;有的寡糖素可使植物產生過敏性死亡,使得病原物不能進一步擴散;還有的寡糖素參與調控植物的形態建成。細胞壁中的伸展蛋白除了作為結構成分外,還有防病抗逆的功能。如黃瓜抗性品種感染一種黴菌后,其細胞壁中羥脯氨酸的含量比敏感品種增加得快。
4、其他功能細胞壁中的酶類廣泛參與細胞壁高分子的合成、轉移、水解、細胞外物質輸送到細胞內以及防禦作用等。
5、參與細胞間的相互粘連,即“胞間連絲”。
研究發現,細胞壁還參與了植物與根瘤菌共生固氮的相互識別作用,此外,細胞壁中的多聚半乳糖醛酸酶和凝集素還可能參與了砧木和接穗嫁接過程中的識別反應。應當指出的是,並非所有細胞的細胞壁都具有上述功能,每一類細胞的細胞壁功能都是由其特定的組成和結構決定的。
基本信息
- 中文名
- 細胞壁
- 外文名
- cell wall
- 研究對象
- 植物、真菌、藻類和原核生物
- 運用領域
- 生物、科技
- 拼音
- xì bāo bì
- 術語類別
- 生物術語
- 出現時間
- 大約在35億年前
- 位置
- 細胞膜之外緊挨著細胞壁