群體遺傳學
群體遺傳學
群體遺傳學是研究群體的遺傳結構及其變化規律的學科,研究群體中基因的分佈,基因頻率和基因型頻率的維持和變化。其研究對象是生物群體,這裡的群體是指孟德爾氏群體。它是由同一物種組成的較大的有性繁殖群體。群體內的個體間是隨機交配的,遵循孟德爾遺傳規律。醫學研究群體遺傳是要探討遺傳病的發病頻率,遺傳方式及其基因頻率和變化的規律,從而了解遺傳病在群體中的發生和散布的規律,為預防、監測和治療遺傳病提供重要的信息和措施。(李英碧)
群體遺傳學最早起源於英國數學家哈迪和德國醫學家溫伯格於1908年提出的遺傳平衡定律。以後,英國數學家費希爾、遺傳學家霍爾丹(Haldane JBS)和美國遺傳學家賴特(Wright S)等建立了群體遺傳學的數學基礎及相關計算方法,從而初步形成了群體遺傳學理論體系,群體遺傳學也逐步發展成為一門獨立的學科。群體遺傳學是研究生物群體的遺傳結構和遺傳結構變化規律的科學,它應用數學和統計學的原理和方法研究生物群體中基因頻率和基因型頻率的變化,以及影響這些變化的環境選擇效應、遺傳突變作用、遷移及遺傳漂變等因素與遺傳結構的關係,由此來探討生物進化的機制並為育種工作提供理論基礎。從某種意義上來說,生物進化就是群體遺傳結構持續變化和演變的過程,因此群體遺傳學理論在生物進化機制特別是種內進化機制的研究中有著重要作用。
在20世紀60年代以前,群體遺傳學主要還只涉及到群體遺傳結構短期的變化,這是由於人們的壽命與進化時間相比極為短暫,以至於沒有辦法探測經過長期進化后群體遺傳的遺傳變化或者基因的進化變異,只好簡單地用短期變化的延續來推測長期進化的過程。而利用大分子序列特別是DNA序列變異來進行群體遺傳學研究后,人們可以從數量上精確地推知群體的進化演變,並可檢驗以往關於長期進化或遺傳系統穩定性推論的可靠程度。同時,對生物群體中同源大分子序列變異式樣的研究也使人們開始重新審視達爾文的以“自然選擇”為核心的生物進化學說。20世紀60年代末、70年代初,Kimura、King和Jukes相繼提出了中性突變的隨機漂變學說:認為多數大分子的進化變異是選擇性中性突變隨機固定的結果。此後,分子進化的中性學說得到進一步完善,如Ohno關於複製在進化中的作用假說:認為進化的發生主要是重複基因獲得了新的功能,自然選擇只不過是保持基因原有功能的機制;Britten甚至推斷幾乎所有的人類基因都來自於古老的複製事件。儘管中性學說也存在理論和實驗方法的缺陷,但是它為分子進化的非中性檢測提供了必要的理論基礎。“選擇學說”和“中性進化學說”仍然是分子群體遺傳學界討論的焦點。
研究進展

Kreitman
群體遺傳學的研究基礎是DNA序列變異。同源DNA序列的遺傳分化程度是衡量群體遺傳結構的主要指標,其分化式樣則是理解群體遺傳結構產生和維持的進化內在驅動力諸如遺傳突變、重組、基因轉換的前提。隨著DNA測序越來越快捷便利及分子生物學技術的飛速發展,越來越多的全基因組序列或者基因序列的測序結果被發表,基因在物種或群體中的DNA多態性式樣也越來越多地被闡明。
植物中,對擬南芥和玉米基因組的DNA多態性的調查最為系統,研究報道也較多。例如,Nordborg等對96個樣本組成的擬南芥群體中的876個同源基因片段(0.48Mbp)的序列單核苷酸多態性進行了調查,共檢測到17000多個SNP,大約平均每30bp就存在1個SNP位點。而Schmid等的研究結果顯示:擬南芥基因組核甘酸多態性平均為0.007(W)。Tenaillon等對22個玉米植株的1號染色體上21個基因共14420bp序列的分析結果顯示玉米具有較高的DNA多態性(1SNP/27.6bp、=0.0096)。Ching等研究顯示:36份玉米優系的18個基因位點的非編碼區平均核苷酸多態性為1SNP/31bp,編碼區平均為1SNP/124bp,位點缺失和插入則主要出現在非編碼區。此外,其他物種如向日葵、馬鈴薯(Solanum tuberosum)、高粱、火矩松(Pinus taeda L.)、花旗松(Douglas fir)等中部分基因位點的DNA多態性也得到調查,結果表明不同的物種的DNA多態性存在較大的差異。
群體指的是孟德爾群體,即一群相互繁育的個體。一個最大的孟德爾群體是一個物種。一個群體中全部個體所共有的全部基因稱為基因庫。群體中各種基因的頻率,以及由不同的交配體制所帶來的各種基因型在數量上的分佈稱為群體的遺傳結構。獲知了不同世代中遺傳結構的演變方式就可探討生物的進化過程並據以培育各種新的生物品系和品種。可是群體遺傳學並不等同於進化遺傳學。後者探討的是物種內變異轉化為物種間變異的過程,即物種的形成和絕滅,而前者則僅僅涉及品系間、品種間和亞種間等的變遷。
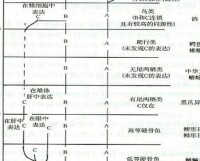
群體遺傳學機制

群體遺傳學規律
基本信息
- 中文名
- 群體遺傳學
- 外文名
- population genetics
- 類型
- 遺傳學分支學科
- 時間
- 1908年後
- 起源
- 遺傳平衡定律