神經嵴
神經嵴
脊椎動物胚胎髮育中的一種過渡性結構,是在神經管建成時位於神經管和表皮之間的一條縱向的細胞帶。1868年德國胚胎學家W.希思首次在雞胚描述了這一構造。

神經嵴
使脊椎與其他動物如此不同的很多特點都與頭有關,其中包括頭蓋骨和專門化的眼睛及耳朵。頭的形式在很大程度上是動物由神經嵴(神經冠)細胞決定的,這些細胞源自胚胎中被稱為“神經板”的、形成中樞神經系統的區域,然後在整個身體中遷移,幫助形成各種不同組織。神經嵴的起源一直是一個謎。這項研究顯示,作為脊椎動物的一個遠親,獨居海鞘有一個源自“神經板”邊緣的細胞系。這些細胞表達幾個神經嵴分化基因,但卻沒有神經嵴的某些明確特性(如長距離遷移)。然而,調控基因Twist的定向“誤表達”就足以誘導這些特性中的其中一些。這些結果表明,神經嵴基因網路大部分是在被囊動物和脊椎動物分化前出現的,“間葉細胞”決定因子(如Twist)向“神經板”外胚層中的“共擇”,對於脊椎動物“新頭”的出現極為關鍵。
形成
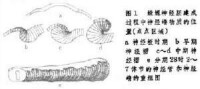
神經嵴
各種脊椎動物的神經嵴細胞的發育命運和遷移行為極為相似,但同一胚胎不同部位的嵴細胞的最終定位和分化卻有很大差別。
在胚胎髮育過程中,神經嵴的形成及其細胞遷移表現明顯的自前而後的順序。因此,在某一發育階段,同一胚胎前後不同部位的神經嵴形成和細胞遷移處於不同的階段。例如雞胚發育第2天,神細褶剛剛在胚體後端融合時,頭部神經嵴已形成並開始細胞遷移,中段神經嵴雖然形成但細胞遷移尚未開始。又如在蛙類,頭部神經嵴細胞遷移開始於神經褶形成時,但軀幹部嵴細胞的遷移直到神經管關閉或閉合以後才開始。
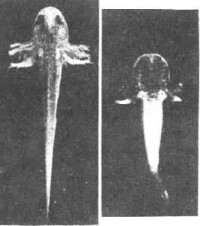
切片遷移 神經嵴細胞的遷移可以根據嵴細胞的顯著形態學特徵的連續切片上進行觀察;也可用移植、活體染色和同位素示蹤的方法追蹤嵴細胞的遷移、定位和最終分化命運。在神經褶形成時或神經管形成之後,頭部的嵴細胞向側腹方遷移,它們在咽囊與咽囊之間結隊而行,形同一股股的細胞流。遷移到第1和第2咽囊之間的細胞,形成舌弓;再后的細胞形成1~4鰓弓;眼前方的嵴細胞則參與顱骨的形成(圖2)。頭部的神細嵴細胞還形成色素細胞和頭部間葉細胞,一部分腦神經節(包括Ⅴ、Ⅶ、Ⅷ、Ⅸ、Ⅹ)也是由神經嵴細胞和頭部外胚層增厚的基板混合組成的。兩棲類的齒乳突也由神經嵴細胞發育而成。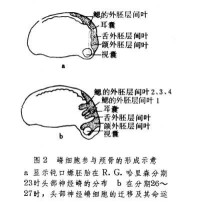 軀幹部的神經嵴細胞主要參與周圍神經系統的形成,併發育為色素細胞。從圖3可以看出,當神經嵴在神經管背方形成之後,嵴細胞便沿著神經管向左右分開。一部分嵴細胞沿著神經管和體節的間隙向腹方遷移,其中一些細胞停留在神經管(脊髓)兩側聚積成緻密的細胞團,成為脊神經節的原基;另一些繼續移行到背主動脈兩側形成交感神經節的原基。神經嵴細胞也參與賢上腺髓部的建造。軀幹部神經嵴細胞的另一個發育方向是形成各種色素細胞,如黃色素細胞、淺棕色色素細胞、銀色素細胞和黑色素細胞。這些嵴細胞以分散的遷移方式在真皮中遷移到胚胎各處,當時並不顯示色素細胞的特徵,直到相當晚的發育階段才有所表現。20世紀初一些學者根據對魚類和兩棲類的觀察就提出過色素細胞來源於神經嵴的設想,後來美國胚胎學家R.G.哈里森發現組織培養的蛙神經管周圍有帶有色素的細胞,因而想到它們很可能來自神經嵴。德國胚胎學家O.曼戈爾德和J.F.K.霍爾特弗雷特移植或外植預定神經板不僅能產生神經組織而且也產生色素細胞,移植的預定表皮則不發生色素細胞。切除蠑螈神經胚期的神經褶(其中包括神經嵴物質),結果在手術範圍內完全缺乏色素細胞,而離體培養的神經褶卻能產生大量的色素細胞,直接證明了色素細胞起源於神經嵴(圖四)。後來的學者應用類似方法在鳥類和哺乳類也證明色素細胞來源於神經嵴。1980年M.西貝爾-布盧姆等指出,神經嵴細胞的克隆能產生神經原和色素細胞。
軀幹部的神經嵴細胞主要參與周圍神經系統的形成,併發育為色素細胞。從圖3可以看出,當神經嵴在神經管背方形成之後,嵴細胞便沿著神經管向左右分開。一部分嵴細胞沿著神經管和體節的間隙向腹方遷移,其中一些細胞停留在神經管(脊髓)兩側聚積成緻密的細胞團,成為脊神經節的原基;另一些繼續移行到背主動脈兩側形成交感神經節的原基。神經嵴細胞也參與賢上腺髓部的建造。軀幹部神經嵴細胞的另一個發育方向是形成各種色素細胞,如黃色素細胞、淺棕色色素細胞、銀色素細胞和黑色素細胞。這些嵴細胞以分散的遷移方式在真皮中遷移到胚胎各處,當時並不顯示色素細胞的特徵,直到相當晚的發育階段才有所表現。20世紀初一些學者根據對魚類和兩棲類的觀察就提出過色素細胞來源於神經嵴的設想,後來美國胚胎學家R.G.哈里森發現組織培養的蛙神經管周圍有帶有色素的細胞,因而想到它們很可能來自神經嵴。德國胚胎學家O.曼戈爾德和J.F.K.霍爾特弗雷特移植或外植預定神經板不僅能產生神經組織而且也產生色素細胞,移植的預定表皮則不發生色素細胞。切除蠑螈神經胚期的神經褶(其中包括神經嵴物質),結果在手術範圍內完全缺乏色素細胞,而離體培養的神經褶卻能產生大量的色素細胞,直接證明了色素細胞起源於神經嵴(圖四)。後來的學者應用類似方法在鳥類和哺乳類也證明色素細胞來源於神經嵴。1980年M.西貝爾-布盧姆等指出,神經嵴細胞的克隆能產生神經原和色素細胞。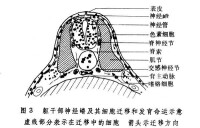
 影響神經嵴細胞遷移、定位及其基因表達的因素神經嵴細胞在明顯表達各自的表型之前就廣泛地遷移並精確地在胚胎各處定位,這一事實曾引起人們極大的興趣並提出許多假想。神經嵴細胞的遷移完全是任意的,還是存在著優先選擇的途徑,如果有優先的通路,是什麼因素在遷移的路上設置地形的界限,在什麼條件下,起初聚集在神經嵴上的細胞開始分散到整個胚胎,什麼時候,在什麼條件下這些細胞就停止遷移並準確地在胚胎定位。事實上,這些問題都是發育的普遍性問題中的一些特定的例子,關係到漸進的決定過程在細胞水平的性質和機制;形態發生過程的細胞基礎和機制,以及細胞基因表達的機制。已經觀察到,神經嵴細胞離開神經管背部后直線地從它們的發源地遷移開,不論神經嵴是處於正常的位置還是在某種不正常的環境中都是如此。例如,即使當神經管與其相連的嵴被背腹顛倒放置於胚胎中,細胞遷移的方向與神經管的背腹方位仍能保持正常的相關,仍然有兩條細胞流,以神經管為基準大都是一條向腹面、一條向側面。所以早期神經嵴細胞遷移的定向像是以某種方式依賴於神經管的。也許是由於神經管以某種形式的“接觸引導”造成的。另一方面,早期嵴細胞遷移的最基本的趨勢是離開它們的發源地而不管它們的環境方位。因此,遷移的方向看來是不依賴細胞遷移中的組織環境的。對於嵴細胞的遷移現象已有一些假說性的解釋,例如化學濃度等級說,接觸抑制說以及細胞與細胞之間相互親和力改變說等,但都沒有足夠的證據。分節遷移神經嵴細胞的分節遷移現象與中胚層組織的存在狀態有關。例如,人為地除去生肌節或使生肌節排列紊亂,則脊神經節不能按節排列。在離體培養條件下,神經嵴細胞在均勻的基質中均勻地散開。當把以3 H-胸腺嘧啶核苷標記的神經管(連同神經嵴)在原位倒置時,遷移入正常分節的肌節間葉細胞中的標記嵴細胞呈分節分佈。如果把標記的神經管移植到側部不分節的間葉細胞中,標記嵴細胞則呈分散狀分佈。這些都表明,分節的中胚層構造決定神經嵴細胞的分節分佈。當然,也不能忽視神經管可能起某種促進作用。如在標記神經嵴細胞遷移到肌節間葉之後,再將肌節間葉(連同遷入的嵴細胞)移植到絨毛尿囊膜上,標記的嵴細胞保持分散狀態而不聚集;如果在做上述手術時,連同神經管一起移植到絨毛尿囊膜上,則標記的嵴細胞聚合形成神經節。從神經嵴細胞遷移的順序及其未來的分化命運來看,形成腸壁色素細胞和交感神經節神經母細胞的嵴細胞最早遷移,而後來遷出的嵴細胞大都定位在神經管兩側肌節間葉中,併發育為脊神經節(一部分形成神經鞘細胞)。神經嵴細胞的基因表達在很大程度上依賴於其周圍的環境條件。例如,把頭部神經褶(包括神經嵴細胞)和前腸內胚層一起培養能發育成軟骨,而單獨培養或與其他組織(包括神經板、脊索、中腸或側部中胚層)混合培養則不能形成軟骨。在整體條件下,鰓軟骨的排列、數目和大小也都與咽囊內胚層的存在狀態相關連。這表明前腸內胚層對頭部神經嵴細胞基因的表達有重要作用。鵪鶉胚和雞胚的移植實驗更進一步證明神經嵴細胞最終到達的環境對它們基因表達的重要影響。已經知道,通往腸道的副交感神經節的神經母細胞和腎上腺髓部的嗜酪細胞合成的神經遞質不同,前者合成膽鹼能的而後者合成腎上腺能的。如果把鵪鶉神經管的預定腎上腺區段移植到雞胚預定腸區的副交感神經節部位,根據鵪鶉胚和雞胚細胞核染色反應明顯的差別,可以判定移植到腸區的預定腎上腺能細胞將分化為副交感神經節細胞而且表現膽鹼能神經原的性質。對於神經嵴的發生,嵴細胞的遷移、定位和正常發育命運已有詳細的形態描述和實驗分析,並已初步提示嵴細胞發育的多潛能性以及胚胎環境對嵴細胞遷移、定位和表型表達的影響。然而,我們對單個嵴細胞的發育潛能了解甚少,也不能確切地說明嵴細胞及其後代在何時、何種條件下發生穩定的決定性變化。正在發展的細胞克隆技術將有助於上述問題的分析。如果能使來源於神經嵴細胞的姐妹亞克隆在各種培養條件下表達各種神經嵴細胞的表型,我們將會得到有關神經嵴細胞多潛能性及環境在促進嵴細胞分化方面特殊作用的明確證據。
影響神經嵴細胞遷移、定位及其基因表達的因素神經嵴細胞在明顯表達各自的表型之前就廣泛地遷移並精確地在胚胎各處定位,這一事實曾引起人們極大的興趣並提出許多假想。神經嵴細胞的遷移完全是任意的,還是存在著優先選擇的途徑,如果有優先的通路,是什麼因素在遷移的路上設置地形的界限,在什麼條件下,起初聚集在神經嵴上的細胞開始分散到整個胚胎,什麼時候,在什麼條件下這些細胞就停止遷移並準確地在胚胎定位。事實上,這些問題都是發育的普遍性問題中的一些特定的例子,關係到漸進的決定過程在細胞水平的性質和機制;形態發生過程的細胞基礎和機制,以及細胞基因表達的機制。已經觀察到,神經嵴細胞離開神經管背部后直線地從它們的發源地遷移開,不論神經嵴是處於正常的位置還是在某種不正常的環境中都是如此。例如,即使當神經管與其相連的嵴被背腹顛倒放置於胚胎中,細胞遷移的方向與神經管的背腹方位仍能保持正常的相關,仍然有兩條細胞流,以神經管為基準大都是一條向腹面、一條向側面。所以早期神經嵴細胞遷移的定向像是以某種方式依賴於神經管的。也許是由於神經管以某種形式的“接觸引導”造成的。另一方面,早期嵴細胞遷移的最基本的趨勢是離開它們的發源地而不管它們的環境方位。因此,遷移的方向看來是不依賴細胞遷移中的組織環境的。對於嵴細胞的遷移現象已有一些假說性的解釋,例如化學濃度等級說,接觸抑制說以及細胞與細胞之間相互親和力改變說等,但都沒有足夠的證據。分節遷移神經嵴細胞的分節遷移現象與中胚層組織的存在狀態有關。例如,人為地除去生肌節或使生肌節排列紊亂,則脊神經節不能按節排列。在離體培養條件下,神經嵴細胞在均勻的基質中均勻地散開。當把以3 H-胸腺嘧啶核苷標記的神經管(連同神經嵴)在原位倒置時,遷移入正常分節的肌節間葉細胞中的標記嵴細胞呈分節分佈。如果把標記的神經管移植到側部不分節的間葉細胞中,標記嵴細胞則呈分散狀分佈。這些都表明,分節的中胚層構造決定神經嵴細胞的分節分佈。當然,也不能忽視神經管可能起某種促進作用。如在標記神經嵴細胞遷移到肌節間葉之後,再將肌節間葉(連同遷入的嵴細胞)移植到絨毛尿囊膜上,標記的嵴細胞保持分散狀態而不聚集;如果在做上述手術時,連同神經管一起移植到絨毛尿囊膜上,則標記的嵴細胞聚合形成神經節。從神經嵴細胞遷移的順序及其未來的分化命運來看,形成腸壁色素細胞和交感神經節神經母細胞的嵴細胞最早遷移,而後來遷出的嵴細胞大都定位在神經管兩側肌節間葉中,併發育為脊神經節(一部分形成神經鞘細胞)。神經嵴細胞的基因表達在很大程度上依賴於其周圍的環境條件。例如,把頭部神經褶(包括神經嵴細胞)和前腸內胚層一起培養能發育成軟骨,而單獨培養或與其他組織(包括神經板、脊索、中腸或側部中胚層)混合培養則不能形成軟骨。在整體條件下,鰓軟骨的排列、數目和大小也都與咽囊內胚層的存在狀態相關連。這表明前腸內胚層對頭部神經嵴細胞基因的表達有重要作用。鵪鶉胚和雞胚的移植實驗更進一步證明神經嵴細胞最終到達的環境對它們基因表達的重要影響。已經知道,通往腸道的副交感神經節的神經母細胞和腎上腺髓部的嗜酪細胞合成的神經遞質不同,前者合成膽鹼能的而後者合成腎上腺能的。如果把鵪鶉神經管的預定腎上腺區段移植到雞胚預定腸區的副交感神經節部位,根據鵪鶉胚和雞胚細胞核染色反應明顯的差別,可以判定移植到腸區的預定腎上腺能細胞將分化為副交感神經節細胞而且表現膽鹼能神經原的性質。對於神經嵴的發生,嵴細胞的遷移、定位和正常發育命運已有詳細的形態描述和實驗分析,並已初步提示嵴細胞發育的多潛能性以及胚胎環境對嵴細胞遷移、定位和表型表達的影響。然而,我們對單個嵴細胞的發育潛能了解甚少,也不能確切地說明嵴細胞及其後代在何時、何種條件下發生穩定的決定性變化。正在發展的細胞克隆技術將有助於上述問題的分析。如果能使來源於神經嵴細胞的姐妹亞克隆在各種培養條件下表達各種神經嵴細胞的表型,我們將會得到有關神經嵴細胞多潛能性及環境在促進嵴細胞分化方面特殊作用的明確證據。
神經嵴
神經嵴
神經嵴