向光性
植物生長器官受單方向光照射而引起生長彎曲的現象
植物生長器官受單方向光照射而引起生長彎曲的現象稱為向光性。對高等植物而言,向光性主要指植物地上部分莖葉的正向光性。以前認為根沒有向光性反應,然而近年來以擬南芥為研究材料,發現根有負向光性。
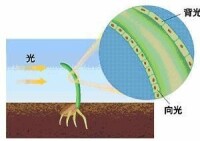
植物的向光性
達爾文根據實驗提出,胚芽鞘尖端受單側光刺激后,就會向下面的伸長區傳遞某種“影響”,造成伸長區背光面比向光面生長快,因而會有胚芽鞘向光性彎曲。
1910年,鮑森詹森的實驗證明,胚芽鞘尖端產生的影響可以透過瓊脂片傳遞給下端。
1914年拜爾的實驗證明,初步證明“影響”是一種化學物質。
1928年荷蘭科學家溫特的實驗證明了“影響”確實是一種化學物質。並命名為生長素。
1931年科學家首次提取成功生長素-吲哚乙酸
實例
植物的向光性以嫩莖尖、胚芽鞘和暗處生長的幼苗最為敏感。生長旺盛的向日葵、棉花等植物的莖端還能隨太陽而轉動。燕麥、小麥、玉米等禾本科植物的黃化苗以及豌豆、向日葵的上下胚軸,都常用作向光性的研究材料。向光性是植物的一種生態反應,如莖葉的向光性,能使葉子盡量處於吸收光能的最適位置進行光合作用。
植物的上部分受光照影響引起生長趨向於光照一側的現象。
王忠(1999)用透明容器(如玻璃缸)水培剛萌發的水稻等,並以單側光照射根,也觀察到根具有負向光性,即種子根向背光的一面傾斜生長(與水平面夾角約60°)。
實驗與研究表明,根具有負向光性,且負向光性與向重性的控制機構相互獨立存在。(石黑和岡田,1994)
植物器官與射來的光線垂直的特性。
傳統的觀點認為,植物的向光性反應是由於生長素濃度的差異分佈而引起的。溫特(1928)用生物測定法顯示生長素活性的分佈比率為向光面32%,背光面68%(相對比值為27∶57)。這是喬羅尼-溫特(Cholodny-Went,1928)假說的主要依據。這個假說認為,植物向光性是由於光照下生長素自頂端向背光側運輸,背光側的生長素濃度高於向光側,使背側生長較快而導致莖葉向光彎曲的緣故。
20世紀70年代,有人分別採用生物測定法和物理化學方法重複了溫特的實驗,用生物測定法得到了與溫特類似的數據,但物理化學方法顯示,向光側和背光側的生長素含量沒有明顯差異。這使人推測,溫特採用的生物測定法由於專一性差,所測出瓊脂塊中的刺激生長的物質可能不單純是IAA,還可能包括生長抑制物質。
對向光性起主要作用的光是420~480nm的藍光,其峰值在445nm左右,其次是360~380nm紫外光,峰值約在370nm。從作用光譜推測,其光敏受體為藍光受體。
以綠色向日葵為材料的測定結果指出,IAA在下胚軸兩側的含量相同,但抑制物質黃質醛(xanthoxin)則是向光側含量高;此後從蘿蔔苗下胚軸中分離與鑒定出蘿蔔寧(raphanusanin)和蘿蔔醯胺(raphanusamide),用蘿蔔寧單側處理可導致黃化蘿蔔苗下胚軸生長失衡,處理側生長受抑;從玉米胚芽鞘中分離與鑒定出6-甲氧基-2-苯並噻唑啉酮(6-?methoxy-2-benzoxazolinone,MBOA)等生長抑制物質,並發現在玉米胚芽鞘中向光側的MBOA含量較背光側高1.5倍,而向光側與背光側IAA含量無明顯差異;外施MBOA或類似物,能導致胚芽鞘發生類似向光彎麴生長的現象,處理側生長慢。另外,還發現這些抑製劑的濃度不僅在向光側增加,而且與光強呈正相關。由此表明,向光性反應並非是背光側IAA含量大於向光側所致,而是由於向光側的生長抑制物質多於背光側,向光側的生長受到抑制的緣故。生長抑製劑抑制生長的原因可能是妨礙了IAA與IAA受體結合,減少IAA誘導與生長有關的mRNA的轉錄和蛋白質的合成。還有試驗表明,生長抑制物質能阻止表皮細胞中微管的排列,引起器官的不均衡生長。
向日葵的生長素主要在莖尖形成,並向基部運輸。生長素的分佈受到光的影響:向光的一側生長素濃度低,背光的一側濃度高。這樣,向光的一側生長區生長較慢,背光的一側生長區生長較快,由此莖就產生了向光性彎曲。向日葵的莖的生長區內含有較高濃度的葉黃氧化素。這種物質與生長素相反,會抑制細胞生長。科學實驗表明:當光由一側照射30分鐘后,在向日葵幼苗生長區的兩側,其葉黃氧化素的濃度分佈為:向光的一側濃度高,背光的一側濃度低。這正好與生長素的濃度分佈規律相反。所以,向日葵的向光性是生長素與葉黃氧化素共同作用的結果。
植物的向光性(phototropism)是植物向性運動(tropicmovement)之一。植物器官的向光性又分為正向光性、負向光性、橫向光性(指器官與射來的光線垂直的特性),但一般植物的莖向光彎曲,呈正向光性。實驗證明,正向光性運動與生長素分佈有關,高級中學《生物》(全一冊)中對此作了說明:光線能使生長素在背光一側比向光一側分佈多,因此,背光側比向光側生長得快,……使植物的莖顯示出向光性。但是,光是如何造成生長素分佈不均勻的,在一些教學輔導書及高校教材中對此說法各異,至今尚無定論。目前主要有以下幾種觀點:
側向運輸觀點
在單側光作用下,生長素自莖的頂端向背光面側向沿薄壁組織向下運輸,背光側的生長素濃度高於向光側,背光側的細胞生長大於向光的一側,這一生長差異引起向光彎曲。溫特(1928年)證明燕麥胚芽鞘經單側照光后,背光一側胚芽鞘尖擴散到瓊脂的生長素含量差不多等於向光一側的2倍。並推測,生長素移動是由於單向光導致胚芽鞘尖不同部位產生電勢差,向光的一側帶負電荷,背光的一側帶正電荷,吲哚乙酸呈弱酸性,其陰離子向背光一側移動,使背光側生長素含量高,生長較快,植株就呈現向光性生長。但這一途徑至今還沒有被證實。
橫向轉移觀點
W.Briggs和他的同事們用玉米胚芽鞘做實驗,結果說明,在單方向照光下,生長素從照光一側向背光側移動。換言之,單側光對生長素的分佈影響就在於能促進生長素沿薄壁組織從向光一側轉移到背光一側,結果背光側的生長素比向光側的多。
生長抑製劑作用觀點
Hasegawa等(1975年)提出了不同的看法。他們重複了溫特的試驗,結果證明,用生物測定法測出燕麥胚芽鞘背光一側擴散的生長素確是多於向光一側的,兩者相對活性與溫特的數據相同,但用物理化學法測定兩側擴散的生長素含量並沒有區別。經過層析分析,發現瓊脂塊中至少有兩種抑制物,向光一側的抑制物活性高於背光一側的。因此,他們認為,燕麥胚芽鞘在單側光照射後向光彎曲,並不是因背光一側生長素含量大於向光一側,而是由於向光一側的生長抑制物比背光一側的多,造成生長速度差異而引起向光彎曲。近年來,一些科學家提出,這些抑制物質主要是蘿蔔寧、蘿蔔醯胺、黃質醛等。
其他觀點
也有人認為,向光性產生的原因是:在單側光刺激下,生長素在向光側和背光側的不對稱運輸及向光側生長抑制物質活性增強兩種現象同時存在,造成了莖的向光側生長較慢。也就是說,植物的向光性不純粹是生長素的作用,而是生長促進物和生長抑制物兩種化合物作用的總反應。LamS.L和A.C.Leopld通過研究向日葵綠色幼苗的向光性后提出了綠葉調節向光性的另外一種見解,他們認為,在照光的和遮蔭的葉片中生長素以及生長素運輸抑製劑的實驗表明,在綠色幼苗中向光彎曲主要是由葉片提供的極性運輸的生長素所調節的,而不需要側向的生長素運輸。
上述幾種觀點都能在一定程度上對植物的向光性作出解釋,但都缺乏更有說服力的證據。目前,向光性產生的機理仍在研究之中,隨著科學的發展和科技手段的現代化,這一疑難問題將會得到科學而圓滿的解釋。

基本信息
- 釋義
- 植物受單方向光照射引起生長彎曲
- 作用
- 獲得更大面積、更多的光照
- 分類
- 正向光性 負向光性
- 泛指
- 植物地上部分莖葉的正向光性
- 中文名
- 向光性
- 外文名
- phototropism