植硅體
植物身體里的結石
植硅體的研究歷史可以簡單地劃分為4個階段:
(1)發現和探索階段(1835-約1900年):植硅體首先在現代植物中被觀察到,並被命名以及從一些環境中分離出來。
(3)生態學研究階段(1955-約1975年):西方土壤學家和植物學家首先開始把植硅體分析應用於古植物學的幾個重要問題,禾本科植硅體的形態得到詳細研究,落葉和針葉植物的硅質體研究也已開始,植硅體分析第一次被應用於研究北美的植被歷史。
(4)現在考古植硅體研究階段(約1971-至今):考古植物學家開始系統地研究植硅體在現代植物中的生產和形態,以及植硅體在考古和地質沉積物中的出現頻度和分佈。
(1)植硅體產量大,分佈範圍廣。從高山到平原,從湖泊到海洋,從森林到草原,凡有植被的地方,即有植硅體的存在。植硅體的這一優點為其研究和應用提供了方便。
(2)植硅體理化性質穩定,具有耐腐蝕性。當植物死亡或凋謝之後,植硅體不會分解,而完整地長期保存於土層中。
(3)植硅體具有極佳的耐高溫特性。在溫度低於700℃時,其物理、化學性質不會發生變化。一般說來,經過高溫處理的考古樣品,往往不再含有化石,因而這種抗高溫性能可使植硅體理想地應用於經高溫處理的依存分析之中。
(4)植硅體形成於植物器官,隨腐爛了的植物,沉積於原地土壤中,而非空氣中,所以,多數植硅體記錄具有原地沉積的特點,特別對考古遺址植物歷史的記錄,更為精細和穩定,優勢十分顯著。
植硅體研究在國外開展較早,在我國是近幾十年才開展起來的一門研究古環境古氣候的新興的邊緣學科,儘管如此,現已在考古學及古環境古氣候解釋及植物起源中取得可喜的成果。利用植硅體分析可以提取沖積物中賦存的古植被信息,利用植硅體進行古植被重建、古氣候學研究是當前國際上生物地層、氣候地層研究的一個新興方向。
植硅體在考古學中的應用
考古土壤中的植硅體組合代表了原地腐爛的植物殘留。因此,植硅體組合在某種程度上能反映微觀的環境變化及相關的文化活動。一般來說,由於人類使用植物的活動,考古遺址各層在堆積過程中,可能存在大量植物的莖和葉。莖和葉腐爛后,植硅體——這種硅化細胞和組織能夠長期保存,而且數量較多。姜欽華等通過對石家河文化時期的堆積物進行植硅石分析,統計有重要意義的植硅體形態類型,發現水稻扇形植硅石,稻啞鈴型,水稻穎殼三種對鑒定稻作農業最為重要的三種植硅體類型。鄭雲飛和俞為潔等對河姆渡遺址和羅家角遺址進行植硅體分析發現了起源於水稻葉片運動細胞植硅體,並把兩遺址的植硅體形狀特徵進行比較,發現此兩遺址的稻分別為粳稻和秈稻。Pearsall等利用十字型植硅體、塊狀植硅體鑒定出玉米。
植硅體研究在古環境解釋中的應用
沉積物中植硅體的組合特徵記錄著一個地方不同時期的植被面貌,植被面貌的變化主要受控於氣候和環境的變化,所以根據植硅體組合特徵可以了解該區域的植被變遷,從而推斷該區域的氣候和環境變化。植硅體分析廣泛地應用於重建古氣候、古環境、古植被。
在我國,經過近幾十年的發展,植硅體定量分析也逐步成熟,通過對黃土、泥炭、海洋沉積物、湖泊沉積物以及考古土壤等沉積物中富含的植硅體進行分析,成功地重建了古環境、古氣候,發現氣候冷暖波動規律及一些特殊環境事件。
呂厚遠和劉東生等通過對渭南、寶雞和蘭州剖面進行植硅體分析表明,末次間冰期以來,在黃土高原南部,主要發育森林草原和發育草原,只有在降水量超過800mm以上,在其後最適宜期,才能有短期的森林植被發育。范斌和許世遠等對巢湖南靈鑽孔的植硅體化石進行系統研究,劃分出六個植硅體組合帶,並根據各組合帶反映的古生態、古環境特徵,發現自5600aBP以來氣候分為3個相對寒冷期和3個相對溫暖時期,並泰倫了個時期巢湖流域的環境變遷。顧延生等對浙江長興中-晚更新世紅土進行分析,發現含有豐富的植硅體形態類型,主要來自草本植物、闊葉和裸子植物等。運用植硅體和分子化石的相關指標揭示了紅土沉積時期的植被與氣候信息。
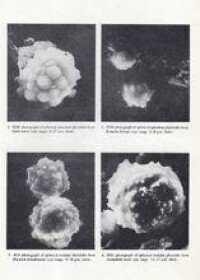
掃描電子顯微鏡下植硅體形態
方型、長方型( Parallelepipedal bulliform cell)、扇型( Cuneifor bulliform cell):從植物細胞學的角度,扇型、方型、長方型植硅體都來源於植物體內的機動細胞,也就是說他們具有一定的同源性,並且,有少量的方型和扇型形態本來就是同一植硅體個體的不同側面,因此,方型、長方型、和扇型的分佈呈現非常相似的側面。扇型呈扇狀或楔形,大小在70μm左右,多產於植物葉片機動細胞,多種植物產此形態,其中禾本科中多產於畫眉草亞科和黍亞科,竹亞科也產扇形,主要分佈在我國的亞熱帶到熱帶地區。
啞鈴型( bilobate):形狀如啞鈴,有雙葉片,大小在25μm左右,主要形成於表皮短細胞,主要產於黍亞科,蘆竹亞科植物。其兩鈴外端輪廓比較薄弱,柄較短;十字型,其兩鈴分叉。多產生於C4植物,主要分佈於熱帶、亞熱帶濕潤地區。
多鈴型( Cylindrical Polylobate):有二個或兩個以上的葉片串狀排列,葉片對稱分佈,大小小於35μm,其分類學意義不明確,有可能產生於黍亞科、畫眉草科亞科、早熟禾亞科。
十字型( Cross):有四個葉片,大小為15μm左右,主要形成於表皮短細胞,多產生於黍亞科,分佈於熱帶、亞熱帶暖濕地區。
短鞍型( Square saddle):形似馬鞍,其面寬大於高,小於15μm,形成於表皮短細胞,多產生於畫眉草亞科,主要分佈在熱帶、亞熱帶乾旱、半乾旱地區,短鞍型也產生於蘆竹亞科,其平面形態如帶柄的水杯。
長鞍型( Oblong concave saddle):長鞍型和竹節型是同一形態不同側面的描述名稱,是相對畫眉草亞科的短鞍型和蘆竹亞科的中鞍型而言的。大小小於20μm,主要形成於表皮短細胞,長鞍型和竹節型是竹亞科的標誌形態,主要分佈於溫暖的半乾旱到濕潤地區,要求有較高的溫度和降水量,多為C3植物。
齒型( Trapeziform sinuate or crenate)、帽型( rondel):齒型分為短齒型和長齒型,短齒型形如平底船或不規則四邊形狀,長齒型兩邊齒狀平行,齒狀或尖、圓或呈直角形,大小在35μm左右;帽型有各種形狀,最基本的形狀象展開的裙子,有的有紋飾突起,大小在15μm左右,主要形成於表皮短細胞;這兩種形態主要形成於早熟禾亞科、畫眉草亞科、蘆竹亞科,早熟禾亞科屬C3植物,主要分佈在溫涼、濕潤的高緯度、高海拔地區。
平滑棒型( Elongate smooth)、突起棒型( Elongate echinate):呈棒狀,大小在100μm左右,無明顯的分類學意義,主要形成於表皮細胞的長細胞,主要分佈在溫帶到亞熱帶地區。
長尖型、短尖型( Unciform hair cell):形狀呈針狀或鉤狀,大小多小於50μm,主要形成於毛狀細胞,主要分佈於溫帶到亞熱帶的地區土壤中。
多面體型( Polyhedron aggregate)、導管型( Vascular tissue)、靴型( Clavate contorted)、薄板型( Platelet polygon)、球型( Globular granulate)等多來自於木本植物。
多邊帽型( Polyhedron with conical projection):鏡下多為空心錐形,其形態表現為一個大圓裡面有個小圓,或是在五邊形或六邊形底盤上中心發育一個圓椎體,大小為40μm,在莎草科植物中居多,多生長於較濕環境。
