Po徠st-transcriptionalGeneSilencing,PTGS。轉錄后基因沉默(PTGS)是近10年發現的一種生物(特別是真核生物)細胞抵抗外來核酸入侵及保持生物自身基因組完整性的防禦機制,特別是與生物的病毒抗性密切相關.PTGS最初在植物內發現,近幾年又分別在真菌、動物等生物細胞內發現。經過10年的研究,我們對PTGS的機制和特點有了相當的了解。這不但對深入地了解基因的表達調控機制意義重大,而且還可為人們如何調控和利用PTGS奠定基礎.
徠研究結果發現有大量的轉基因植株不能正常表達,通常這並不是由於轉基因的缺失或突變引起的,而是基因失活的結果。這種失活的現象稱為基因沉默。部分的植物中的基因沉默是在轉錄后發生的,稱為轉錄后基因沉默。
轉錄后基因沉默
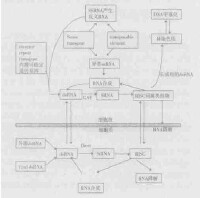
從遺傳分析看, 通過特異性擴增dsRNA 可以提高RNA i介導基因沉默的效率。最近在
線蟲的研究中發現, siRNA 可能是合成 dsRNA 的特殊引物, 在RNA 依賴RNA 聚合酶(RdRP)作用下, 以靶mRNA 為模板合成dsRNA。新生成的dsRNA 在D icer 酶的作用下, 裂解產生新的siRNA , 新生成的siRNA 又可進入上述循環。大量集中的siRNA 可以形成R ISC複合物, 這樣可以提高
mRNA 降解的效率。在這種RNA i過程中, 對靶mRNA 的特異性擴增有助於增強RNA i的特異性基因監視功能, 每個細胞只需少量的dsRNA 就能完全關閉相應基因的表達。由於siRNA 合成的產物和靶mRNA 裂解物極有可能在胞液中, 因而線蟲中依靠RdRP 進行特異性擴增步驟, 可以推測是發生在
細胞質中(圖1) , 當外源注射dsRNA式轉基因時, 引起RNA i則需要RDE21 和RDE24, 但對於較短的反義RNA s 介導的基因沉默則不需要RDE21 和RDE24。通過對網柱原蟲(D icty ostelium )的研究又發現, RNA i過程中需要一種類似RdRP 作用的類似RRPA。RRPA 是具有含N 2末端的RNA 解旋酶結構(在其他生物RdRP s 中還沒發現這種具有
解旋酶活性的結構)。但令人奇怪的是, 線蟲中的D icer酶也具有與RRPA 非常類似的解旋酶的結構域, 這可以說明網柱原蟲的D icer 酶與RRPA 中的這種具有RNA 解旋酶活性結構域可以發生交換。倘若這一點被證實, 則由D icer 酶與RdRP 共同作用產生 dsRNA 引發RNA i, 則就可以直接由D icer 酶作用, 從而更容易快速地獲得dsRNA。類似RdRP s 活性的酶的作用機制還有待進一步證實。當弄清線蟲體內RRF23 (可能的RdRP)作用機制后, 有望得到證實。RRF3 在線蟲某些組織中可以增強RNA i敏感性, 因而RRF3可作為RNA i反應的負面調節物。