變形運動
變形運動
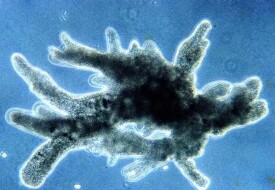
變形運動是細胞的變形運動,這種變形運動在原生動物變形蟲表現得最為典型。
變形運動又稱偽足運動(pseudopodial movement)。除了作為根足蟲類特有的運動形式以外,變形運動是各種變形細胞或遊走細胞的屬性,廣泛地分佈在後生生物界。
變形運動既有把偽足附著在基底上的細胞移動運動(如:變形蟲類,變形菌類的變形體,蛔蟲的精子,脊椎動物的原始生殖細胞,淋巴球,白血球,低等無脊椎動物的排出遊走細胞,成長中的神經纖維等),又有僅在攝食中使遊離性偽足伸縮、屈曲的局部運動(如有孔蟲類,太陽蟲類,脊椎動物的網內皮系細胞,巨噬細胞等)。
組織培養下的細胞以及一般細胞分裂和胚胎髮生時的形態形成運動,都具有變形運動的因素。變形運動方式可因細胞的種類、特別是偽足的樣式不同而異。具有葉狀偽足的變形蟲(特別是蛞蝓變形蟲Amoeba limax和大變形蟲A.proteus)作為整個變形運動的基本型,正在深入進行研究。
體表有稱為質膜(plasma lemma)的薄的細胞膜,質膜之下為流動的透明的外質,外質的內層為粘稠而不流動的凝膠質形成的圓筒狀殼,其內為內質,內質是被圍在殼內流動的溶膠質。溶膠質不斷地流向變形蟲的移動方向,在前端形成偽足。由於質膜的粘著性,偽足對基底的附著,和細胞後邊的凝膠質轉變為溶膠質,細胞前邊的溶膠質,轉變為凝膠質(凝溶轉換)是變形移動運動的二個主要原因。
變形移動運動速度是0.5-4.6微米/秒。絲狀偽足和軸狀偽足運動的形式不同,可以看到前者乃是純粹的原生質流動,後者乃是典型的原生質收縮。從表現變形運動的變形蟲和白血球中可以分離、精製出收縮性蛋白質為肌纖蛋白和肌球蛋白。這些物質的物理化學性質與從兔的橫紋肌和變形菌的變形體中分離的很相似。
肌纖蛋白聚合而成直徑為6納米的F-肌纖蛋白,在細胞中作為微絲存在。可以認為變形蟲的運動與橫紋肌一樣,是由ATP(三磷酸腺苷)和這些收縮性蛋白質的相互作用引起的。在太陽蟲類,放射蟲類的軸(針)狀偽足內,有發達的直徑為25納米的軸絲(偽足軸),它們有很強的雙折射性(復曲折射)。可以認為偽足的構造和收縮性取決於這些軸絲,但是軸絲的構造尚不太清楚。
基本信息
- 中文名
- 變形運動
- 外文名
- amoeboid movement
- 分佈
- 後生生物界
- 所屬學科
- 生物學