微絲
微絲
微絲(microfilaments)是由肌動蛋白分子螺旋狀聚合成的纖絲,又稱肌動蛋白絲(actin filament),與微管和中間纖維共同組成細胞骨架,是一種所有真核細胞中均存在的分子量大約42kDa的蛋白質,也是一種高度保守的蛋白質,因物種差異(例如藻類與人類)的不同不會超過20%。微絲對細胞貼附、鋪展、運動、內吞、細胞分裂等許多細胞功能具有重要作用。
微絲的主要功能有:微絲聚集成束,沿平行於胞質環流的方向排列,控制細胞的胞質環流。花粉管的生長也與微絲有關。
微絲
微絲與它的結合蛋白(binding protein)以及肌球蛋白(myosin)三者構成化學機械系統,利用化學能產生機械運動。由微絲形成的微絲束稱為應力纖維,常橫貫於細胞長軸。脊椎動物肌動蛋白分為α、β和γ三種類型,α型分佈於心肌和橫紋肌細胞中,α及γ型分佈於平滑肌細胞中,β及γ型分佈於非肌細胞中。聚合的及非聚合態的肌動蛋白能與其多種結合蛋白相互作用,這些結合蛋白對肌動蛋白的聚合及對微絲的穩定、長度及分佈具有調節作用。
微絲
微絲首先發現於肌細胞中, 在橫紋肌和心肌細胞中肌動蛋白成束排列組成肌原纖維,具有收縮功能。微絲也廣泛存在於非肌細胞中。在細胞周期的不同階段或細胞流動時,,它們的形態、分佈可以發生變化。因此,非肌細胞的微絲同胞質微管一樣, 在大多數情況下是一種動態結構,以不同的結構形式來適應細胞活動的需要。
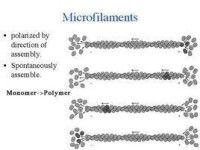
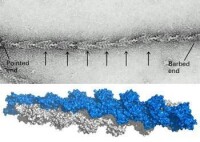
微絲能被組裝和去組裝。當單體上結合的是ATP時,就會有較高的相互親和力,單體趨向於聚合成多聚體,就是組裝。而當ATP水解成ADP后,單體親和力就會下降,多聚體趨向解聚,即是去組裝。高ATP濃度有利於微絲的組裝。所以當將細胞質放入富含ATP的溶液時,細胞質會因為微絲的大量組裝迅速凝固成膠。而微絲的兩端組裝速度並不一樣。快的一端(+極)比慢的一端(-極)快上5到10倍。當ATP濃度達一定臨界值時,可以觀察到+極組裝而-極同時去組裝的現象,被命為“踏車行為”。
微絲
微絲的組裝可用踏車模型(treadmiling model)和非穩態動力學模型(dynamic instability)來解釋,但後者更為合理。ATP是調節微絲組裝的動力學不穩定性行為的主要因素。另外,微絲結合蛋白(actin-binding protein,ABP)對微絲的組裝也有調控作用。
微絲的組裝和去組裝受到細胞質內多種蛋白的調節,這些蛋白能結合到微絲上,影響其組裝去組裝速度,被稱之為微絲結合蛋白(association protein)。
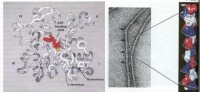
微絲的組裝先需要“核化”(nucleation),即幾個單體首先聚合,其它單體再與之結合成更大的多聚體。Arp複合體(Actin related-protein)是一種能與肌動蛋白結合的蛋白,它起到模板的作用,促進肌動蛋白的多聚化。Arp複合體由Arp2,Arp3和其它5種蛋白構成。
封閉蛋白(end-blocking protein)則是微絲兩端的“帽子”。當這種蛋白結合到微絲上時,微絲的組裝和去組裝就會停止。這對一些長度固定的蛋白來說很重要,如細肌絲。
而前纖維蛋白(Profilin,或譯 G肌動蛋白結合蛋白)則是促進多聚的,相應地促解聚的蛋白則有絲切蛋白(Cofilin)。纖絲切割蛋白(filament severing protein),如溶膠蛋白(Gelsolin),能將微絲從中間切斷。粘著斑蛋白(Vinculin)則能固定微絲到細胞膜上,形成粘著斑。交聯蛋白(cross-linking protein)有兩個以上肌動蛋白結合位點,起到連接微絲的作用,其中,絲束蛋白(fimbrin)幫助微絲結成束狀,而細絲蛋白(filamin)則將微絲交聯成網狀。
微絲除參與形成肌原纖維外還具有以下功能:
微絲
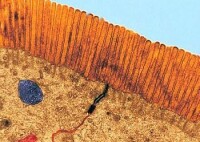
小腸微絨毛
細胞的變形運動中,微絲起著關鍵的作用。過程可分為以下四步:
①微絲纖維生長,使細胞表面突出,形成片足(lamellipodium)。
②在片足與基質接觸的位置形成粘著斑。
③在myosin的作用下微絲纖維滑動,使細胞主體前移。
④解除細胞後方的粘和點。如此不斷循環,細胞向前移動。阿米巴原蟲、白細胞、成纖維細胞都能以這種方式運動。
有絲分裂末期,兩個即將分離的子細胞內產生收縮環,收縮環由平行排列的微絲和myosin II組成
微絲
在精卵結合時,微絲使頂體突出穿入卵子的膠質里,融合后受精卵細胞表面積增大,形成微絨毛,微絲參與形成微絨毛,有利於吸收營養。
如細胞器運動、質膜的流動性、胞質環流均與微絲的活動有關,抑制微絲的藥物(細胞鬆弛素)可增強膜的流動、破壞胞質環流。
肌動蛋白在塑造和維持細胞形態方面扮演著重要的角色,同時也擔負著支撐細胞各項功能的作用,比如細胞移動,細胞分裂,細胞內運輸等。對於神經細胞來說,肌動蛋白更是神經元極性,作用因子運輸,神經突起生長,以及突觸結構穩定必不可少的元件。
可切斷微絲纖維,並結合在微絲末端抑制肌動蛋白加合到微絲纖維上,特異性的抑制微絲功能。
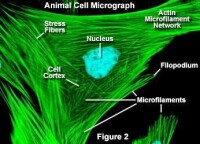
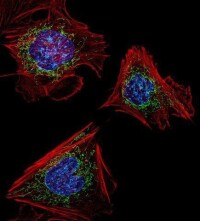
微絲的鬼筆環肽染色

基本信息
- 中文名
- 微絲
- 別名
- 肌動蛋白絲
- 類型
- 生物學名詞
- 界
- 植物界
- 地位
- 對細胞起著很大的作用
- 學名
- microfilaments, actin filament
- 實質
- 分子量大約42kDa的蛋白質