中樞抑制
中樞抑制
在任何反射活動中,中樞內既有興奮活動又有抑制活動。某一反射進行時,某些其他反射即受抑制,例如吞咽時呼吸停止、屈肌反射進行時伸肌即受抑制。反射活動有一定的次序、一定強度,並有一定的適應意義,是反射的協調功能的表現。反射活動所以能協調,就是因為中樞內既有興奮活動又有抑制活動;如果中樞抑制受到破壞,則反射活動就不可能協調。例如,用士的寧破壞脊髓抑制活動后,任何一個微弱刺激會導致四肢出現強烈的痙攣性收縮,失去了反射活動的協調性。根據中樞抑制產生機制的不同,抑制可分為突觸后抑制和突觸前抑制兩類。
在哺乳類動物中,所有的突觸后抑制都是由抑制性中間神經元活動引起的。由這一抑制性神經元發出的軸突末梢釋放的遞質,能使所有與其發生突觸聯繫的其他神經元都發生抑制,都暗生抑制性突觸后電位。按此觀點,一個興奮性神經元通過突觸聯繫能引起其他神經元產生興奮,但不能直接引起其他神經元產生突觸后抑制;它必須首先興奮一個抑制性神經元,轉而抑制其他神經元。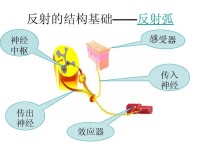
中樞抑制
抑制性突觸后電位脊髓前角運動神經元有的支配伸肌,有的支配屈肌。來自伸肌肌梭的傳入神經衝動,能興奮伸肌運動神經元,也能同時通過抑制性中間神經元轉而抑制屈肌運動神經元。如用微電極插入屈肌運動神經元細胞體內,並刺激伸肌肌梭的傳入神經使屈肌運動神經元發生抑制,可見到其細胞體的突觸后膜出現超極化現象。這時膜電位的數值向-80mV水平靠近。這種超極化膜電位變化稱為抑制性突觸后電位(inhibitory postsynaptic potential,IPSP)。
抑制性突觸后電位的變化與興奮性突觸后電位變化在時程上極相似,但前者為超極化,後者為去極化,變化方向恰相反(圖10-16)。可以設想突觸后膜在超極化狀態下。軸突始段部位將出現內向電流,造成該處不易爆發動作電位,也就表現為抑制。由於這種抑制是突觸后膜出現抑制性突觸后電位所造成的,因此稱為突觸后抑制(postsynaptic inhibition)。抑制性突觸后電位是突觸后膜對CI-通透性增加而形成的。
每組曲線的下線為某一屈肌運動神經元細胞內電位記錄,上線為刺激拮抗伸肌傳入神經時的背根電位記錄。當刺激強度逐步加大時,背根電位逐步增大,超極化電位變化也逐步增大。
突觸后抑制的分類根據抑制性神經元的功能和聯繫方式的不同,突觸后抑制可分為傳入側支性抑制(afferent collateral inhibition)和回返性抑制(recurrent inhibition)。
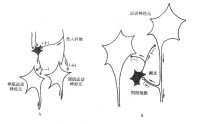
A:傳入側支性抑制;B:回返性抑制
(2)回返性抑制:是指某一中樞的神經元興奮時,其傳出衝動沿軸突外傳,同時又經軸突側支去興奮另一抑制性中間神經元;該抑制性神經元興奮后,其活動經軸突反過來作用於同一中樞的神經元,抑制原先發動興奮的神經元及同一中樞的其他神經元。脊髓前角運動神經元與閏紹細胞之間的聯繫,
就是這種抑制的典型。前角運動神經元發出軸突支配外周的骨骼肌,同時也在脊髓內發出側支興奮閏紹細胞;閏紹細胞是抑制性神經元,其活動經軸突回返作用於脊髓前角運動神經元,抑制原先發動興奮的神經元和其他神經元。這種形式的抑制在海馬和丘腦內也明顯存在。這種抑制是一種負反饋控制形式,它能使神經元的活動及時終止,也促使同一中樞內許多神經元之間的活動能步調一致。丘腦與海馬內許多神經元的活動能夠同步化,就是由於存在回返性抑制環節的緣故。閏紹細胞軸突末梢釋放的遞質是甘氨酸,其作用能被士的寧和破傷風毒素所破壞;在閏紹細胞功能存在後,將出現強烈的痙攣。
前文已述及,軸突可與另一神經元的軸突構成突觸,這種突觸可能是突觸前抑制的結構基礎。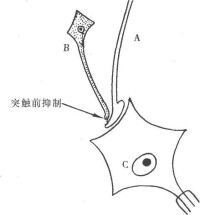
中樞抑制
纖維末梢與運動神經元構成軸突-胞體型突觸,能興奮該運動神經元;b 纖維傳入經過多突觸接替后,末梢與A纖維末梢構成軸突-軸突型突觸,不能直接影響該運動神經元活動。當A纖維興奮傳入衝動抵達末梢時,可引致運動神經元出現興奮性突觸后電位(圖10-18,甲,1);當僅有B纖維興奮衝動傳入時,見不到該運動神經元有反應。如果先使B纖維興奮,一定時間間隔后再使A纖維興奮,則A纖維興奮所引起的興奮性突觸后電位明顯減小(圖10-18,甲,2,2),說明B纖維的活動能抑制A纖維的興奮作用。已知,抵達末梢部位的動作電位是觸發神經遞質釋放的因素,如動作電位大則遞質釋放量大,運動電位小則遞質釋放量小;而動作電位的大小又受到軸突末梢跨膜靜息電位的影響,跨膜靜息電位大則動作電位大,跨膜靜息電位小則動作電位也小。由此認為,突觸前抑制產生的機制是:B纖維傳入經多突觸接替后,興奮抵達末梢交釋放遞質→遞質作用於A纖維末梢使其去極化,從而使末梢跨膜靜息電位變小→A纖維興奮時其末梢的動作電位變小,使釋放的遞質減少→運動神經元的興奮性突觸后電位減小。因此,B纖維的抑制作用是通過使A纖維釋放的興奮性遞質減小而實現的。由於這種抑制是改變了突觸前膜的活動而實現的,因此稱為突觸前抑制。
突觸前抑制在中樞神經系統內廣泛存在,尤其多見於感覺傳入途徑,對調節感覺傳入活動有重要作用。突觸前掏可發生在各類感受器傳入活動之間,也可發生面類感受器的不同感受野活動之間;即一個感覺傳入纖維的興奮衝動進入中樞后,它本身沿特定的傳導路徑向高位中樞,同時通過多個神經元的接替,轉而對其旁的感覺傳入纖維的活動發生突觸前抑制限制其他的感覺傳入活動。由於突觸前抑制產生的潛伏期較長,因此認為傳入神經必須通過兩個以上中間神經元的多突觸接替,才能與其他感覺傳入神經末梢形成軸突-軸突型突觸聯繫。突觸前抑制一般約在刺激傳入神經后20ms左右發展到高峰,而後其抑制作用逐漸減弱,整個抑制過程可持續100-200ms。
在局部神經元迴路中提到了交互性突觸聯繫,這種聯繫可以由一個興奮性突觸和一個抑制性突觸組合而成。在這種聯繫中,一個樹突活動時,首先通過興奮性突觸激活另一個樹突;而後一個權威突活動加強時,卻通過抑制性突觸來抑制前一個樹突的活動,使原行發動興奮的樹突很快受到反饋抑制。這種抑制,發生在局部神經元迴路中,起到了局部的整合作用;而且這種抑制只有樹突的一部分參與活動,不需要整個神經元參與活動。由樹突-樹突型突觸聯繫產生的抑制,稱為樹突-樹突型抑制(dendrodendritic inhibirion),它在視網膜、嗅球、丘腦內都存在,也是中樞抑制的一種形式。
根據抑制性神經元的功能和聯繫方式的不同,突觸前抑制可分為交互抑制和回返性抑制。
交互抑制:
當一個中樞興奮時可同時引起另一中樞發生抑制的現象,稱為交互抑制。例如屈肌反射進行時,屈肌中樞興奮,而伸肌中樞則被抑制。交互抑制是由於一個感覺神經元興奮時,衝動沿傳入纖維進入脊髓后,一方面直接興奮某一中樞的神經元,另一方面經其側枝興奮一個抑制性中間神經元,轉而引起另一中樞的抑制。這種抑制是經傳入神經的側枝而引起的,所以又稱為傳入側枝性抑制。交互抑制可使不同中樞(尤其是功能上拮抗的中樞)之間的活動協調起來;即當一個中樞興奮時,與之拮抗的中樞即發生抑制,兩者互相配合,使反射活動更為協調。
回返性抑制:
一個中樞的興奮活動可通過興奮一個抑制性中間神經元而返回抑制原先發動興奮的神經元的活動,稱為回返性抑制。回返性抑制是由於在反射的傳出途徑中,有抑制性神經元與原先發動興奮的神經元發生環狀聯繫的緣故。這樣,某一中樞的神經元興奮時,一方面經其軸突外傳,另一方面經軸突側枝去興奮一個抑制性中間神經元,由它返回抑制原來神經元的活動,使其活動及時中止。脊髓前角運動神經元與閏紹細胞之間的功能聯繫就是回返性抑制的典型。士的寧和破傷風毒素可破壞閏紹細胞的功能,阻斷了回返性抑制,導致骨骼肌發生痙攣。

基本信息
- 中文名
- 中樞抑制
- 內涵
- 興奮活動、抑制活動
- 分類
- 突觸后抑制和突觸前抑制
- 相關學科
- 生物學