脊索
脊索
脊索(notochord)是身體背部起支持作用的棒狀結構,位於消化道背面、背神經管腹面。在發生上來自胚胎的原腸背壁,后與原腸脫離形成。典型的脊索由富含液泡的脊索細胞組成,外面圍有脊索細胞分泌形成的結締組織鞘,即脊索鞘(notochordalsheath)。脊索鞘常包括內、外兩層,分別為纖維組織鞘和彈性組織鞘。充滿液泡的脊索細胞由於產生膨壓,使脊索既具彈性又有硬度。脊索終生存在低等脊索動物中(如文昌魚)或僅見於幼體時期(如尾索動物)。脊椎動物中的圓口類脊索終身保留,其他類群只在胚胎期出現脊索,後來被脊柱所取代,成體的脊索完全退化或保留殘餘。
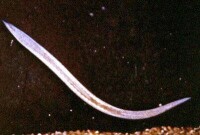
脊索動物
脊索
脊索是一條由多泡狀角質細胞所組成的堅韌、半透明、有彈性的圓柱形不分節結構,其表面覆蓋著數層薄薄的結締組織的脊索鞘(elastica)。脊索是胚胎時期的原腸壁背面加厚外突形成,是內胚層結構。之後顯著伸長和集中,將陷入胚內的外胚層作為襯底,成為身體中心中樞神經系統產生的誘導物質,預定的脊索原基內陷后也明顯地伸長。文昌魚終生具有脊索,圓口類的脊索也不退化而終生存在。脊椎動物的脊索在胚胎期及幼體期具有中軸支持器官的作用,但是其後被周圍結締組織形成的軟骨或硬骨的脊柱所代替,只在脊柱內殘存一些脊索的痕迹。
脊索動物的來源有兩種假說:環節來源說和棘皮來源說。由於棘皮來源說有更多的證據,本文按照棘皮來源說(echinoderm theory)論述。
脊索動物有一些性狀與棘皮動物相同,如后口、三胚層、兩側對稱和分節現象等,說明脊索動物是由棘皮動物進化而來。按照綜合進化論可以推測某些棘皮動物胚胎期變異后體內產生脊索雛形,這種脊索雛形使棘皮動物幼體可以更方便的運動,從而擴大了攝食範圍和增強了逃避敵害的能力,也使得有脊索雛形個體可以到達更遠的地方,降低種內競爭,形成了形成新物種的條件。在自然選擇下體內有脊索雛形的棘皮動物形成了新的物種。雖然由於原始脊索動物沒有化石殘留,但通過比較解剖學和胚胎學的證據,半索動物的幼體──柱頭幼蟲(Torn aria)可能可以作為棘皮動物和脊索動物的過渡型。因為柱頭幼蟲咽喉正中的背側生長的囊狀體從發生學和功用上均類似於脊索,生物學家也稱其為脊索原基。
自然選擇令在運動和支撐身體上有更多優勢的脊索使越來越完善。於是進化出尾索動物(SubphylumUrochordata)。尾索動物仍保留有半索動物的一些特徵:幼體尾部有較為成型的脊索,但在成體內脊索又消失。但是幼體脊索已經開始成型。如現存的海鞘(Ascidiacea)、尾海鞘(Appendiculariae)等生物。
由於脊索不僅能令動物幼體更具生存競爭力,對於成體,脊索也有新的用途,所以變異的那些脊索消失較晚的個體漸漸淘汰了脊索消失早的個體,這些脊索消失較晚的個體又進化出了新的生活方式和身體結構,這就是頭索動物(SubphylumCephalochordata)。從比較胚胎學證據也得出尾索動物進化又產生了頭索動物。頭索動物的脊索縱貫於全身的背部,並終生保留。頭索動物身體呈魚形,體節分明,表皮只有一層細胞,頭部不顯,故稱無頭類(Acrania),如文昌魚。至此,完整的脊索完全產生了。
脊索的出現構成了支撐軀體的主梁,這個主梁使體重有了更好的受力者,體內內臟器官得到有力的支持和保護,運動肌肉獲得堅強的支點,在運動時不致由於肌肉的收縮而使軀體縮短或變形。所有的這些雖然和節肢動物堅硬的幾丁質外鞘作用類似,但是由於脊索在體內,同時比幾丁質外鞘有更好的的韌性,因而使脊索動物身體更靈活,體形有可能向“大型化”發展。這樣,在環境的變化下,脊索動物獲得了新的適應性。
脊索的中軸支撐作用也使動物體更有效地完成定向運動,對於主動捕食及逃避敵害都更為準確、迅捷。脊椎動物頭骨的形成、頜的出現以及椎管對中樞神經的保護,都是在此基礎上進一步完善化的發展。
脊索在發生學和胚胎學上還有誘導周圍細胞分化作用,促進了胚胎期由中胚層發育而來的脊椎和由外胚層發育而來的神經的產生和完善。這一點也是脊索為什麼仍然存在於高等脊索動物的胚胎內的一個原由。
雖然脊索具有一定的彈性和韌性,但它究竟不太堅硬,所能承受的應力是有限的。因此當動物大型化到一定程度時,脊索就不夠用了。之後中胚層靠近脊索的間葉細胞起初集聚變為生骨節(sclerotoma)。在圓口動物如七鰓鰻、盲鰻的脊索附近找到的髓弧片就是進化到這一時期的生骨節產生的脊椎雛形。
隨後生骨節產生的髓弧片開始癒合,直至完全包裹脊索,漸漸出現椎體(centrum)等結構,同時向動物體背側延伸,出現包裹神經管的趨勢。脊索被椎體壓迫,逐漸變細小。但是此時的“脊椎骨”細胞只是失去了原生質的分支,同時細胞間有大量間質,即此時是軟骨脊椎。另外每個脊椎骨環是獨立的。由於是軟骨,此段缺少現存或化石證據,但是在胚胎學的重演律上可以證明,脊椎是先一段一段產生再癒合的。
隨後軟骨開始硬化,每段脊椎(vertebrae)也開始癒合,神經管被脊椎包裹,脊索被分開,形成脊索小球,像一串念珠,夾於脊椎中。鯊魚體內的脊椎就是這樣,頭部還殘留有軟骨和沒封閉完全的骨,脊椎簡單癒合。
之後脊索有不同的去向:
1)脊索小球剛好鑲入兩側的脊椎中央,形成雙凹形(amphicoelous)脊椎。如硬骨魚類
2)脊索小球與後面的脊椎癒合,形成前凹形(procoelous)脊椎。如爬行類和多數無尾類。
3)脊索小球與前面的脊椎癒合,形成后凹形(opisthocoelous)脊椎。如某些兩棲類。
4)脊索小球與兩面都粘連,形成椎間盤(discus intervertebralis),脊椎則形成雙平形(amphiplatyan)。新形成的椎間盤不是脊索,而是由生骨節發生的結構,椎間盤中間很小的髓核才是退化的脊索。如哺乳類。
脊椎已經替代了脊索成為了新的身體支持物,脊椎在靈活性和堅固性兩方面都比脊索要好。從此脊索在高等動物體內消退,變為痕迹器官。
脊索是泡狀細胞組成的棒狀組織,在進化歷程中起到了不可磨滅的作用。最初脊索在低等脊椎動物如無頜類中起著重要的支持作用,為脊索動物提高了生存競爭力。而在其以後的高等脊椎動物中,則被脊椎代替,最後在路生脊椎動物中脊索僅出現在胚胎期。本文重新瀏覽了脊索從出現到消失的歷程,可以發現脊索在脊索動物進化路上的重要性。同時也從側面論述了脊索動物的來源和進化過程中脊索的誘導作用。
基本信息
- 中文名
- 脊索
- 外文名
- notochord
- 拼音
- jǐ suǒ
- 結構組成
- 半透明、堅韌有彈性的無節圓柱形
- 位置
- 位於消化道背面、背神經管腹面
- 所屬系統
- 骨骼系統
- 形態
- 棒狀結構
- 功能
- 身體背部起支持作用
- 組成
- 多泡狀角質細胞
- 保健方法
- 適量的運動
- 性質
- 身體背部起支持作用的棒狀結構
- 發生
- 胚胎的原腸背壁