木質部
木質部
徠木質部是維管植物的運輸組織,負責將根吸收的水分及溶解於水裡面的離子往上運輸,以供其他器官組織使用,另外還具有支持植物體的作用。木質部由導管、管胞、木纖維和木薄壁組織細胞以及木射線組成。木質部一詞是由希臘文 xylon(木材)衍生而來的。木質部中最早形成的部分。一種無穿孔的狹長管狀分子,兩端漸尖,細胞壁明顯增厚,並木質化,成熟后原生質體解體,細胞死亡。
維管植物(蕨類植物、裸子植物和被子植物)中,主要起輸導水分和無機鹽,並有支持植物體作用的複合組織。木質部常與韌皮部結合在一起,在植物體中構成連續的維管系統。木質部一詞是由希臘文 xylon(木材)衍生而來的。
徠為一串高度特化的管狀細胞,其細胞端壁由穿孔相互銜接,其中每一個細胞稱為一個導管分子或導管節。導管在植物體中,主要起輸導水分和無機鹽的作用。在被子植物中,除少數科屬(如昆欄屬Trochodendron,水青樹屬Tetracentron和Winteraceae)外,均有導管;導管也存在於某些蕨類(如卷柏 Sela-ginella,歐洲蕨Pteridium aquilinum)和裸子植物的買麻藤目(Gn
etales)中。導管分子在發育初期是生活的細胞,成熟后,原生質體解體,細胞死亡。在成熟過程中,細胞壁木質化並具有環紋、螺紋、梯紋、網紋和孔紋等不同形式的次生加厚。在兩個相鄰導管分子之間的端壁,溶解后形成穿孔板。只有一個穿孔的,稱為單穿孔板,有多個穿孔的,稱為復穿孔板。復穿孔板的穿孔為長形平行排列的,稱為梯狀穿孔板,成網狀的,稱網狀穿孔板。在導管分子側壁上有簡化的具緣紋孔並排列成各種形式。相鄰導管分子之間的側壁上常有具緣紋孔對。在導管分子與木纖維或木薄壁組織細胞之間有半具緣紋孔對。
導管存在於木質部中,是運輸水分和無機鹽的通道。莖里的導管與根、葉中的導管相通,水分和無機鹽在導管中能自下而上地向枝端運輸。
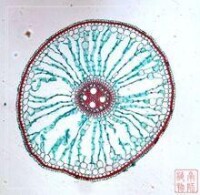
木質部
木纖維是長形,兩端漸尖的厚壁組織細胞,一般具木質化的次生壁,壁常厚於管胞壁。在植物體中主要起支持作用。木纖維可分為纖維管胞和韌型纖維兩種類型。前者為纖維狀管胞,一般具有加厚的次生壁,壁上具雙凸鏡形或裂縫狀開口的具緣紋孔。後者比前者長而壁厚,壁上有單紋孔。這兩種纖維均可具分隔。分隔木纖維廣泛存在於雙子葉植物中,並在成熟的邊材中,殘留有原生質體,起貯藏營養物質的作用。
在次生木質部中一般可分為軸向薄壁組織細胞和徑向薄壁組織細胞(即“木射線”)兩種。其細胞成熟后,一般都具有加厚的次生壁,並木質化。如有次生壁,則厚壁組織細胞壁之間的紋孔對有具緣、半具緣的類型。木薄壁組織細胞主要起貯藏作用。在木材橫切面上,依據軸向薄壁組織細胞與導管相對位置的不同,可分為離管薄壁組織和傍管薄壁組織兩大類。
位於形成層以內次生木質部中的維管射線。其主要功能是橫向運輸和貯藏。在橫切面上,木射線呈輻射狀排列。在切向切面上,射線呈縱線或紡錘形。在徑向切面上,射線呈不同高度的線狀或片狀排列。木射線單列或多列、高度從一層到多層細胞。小的射線也可聚合成群,形成聚合射線。組成木射線的薄壁組織細胞,依據細胞最長軸方向的不同,可分為徑向伸長的橫卧細胞,以及方形或沿縱向伸長的直立細胞兩種類型。如果由同一種類型細胞組成的射線,或僅由射線薄壁組織細胞組成的,稱為同型射線,由不同類型的薄壁組織細胞組成的射線(雙子葉植物)或在裸子植物中由木薄壁組織細胞和射線管胞構成的射線,稱異型射線。
木質部
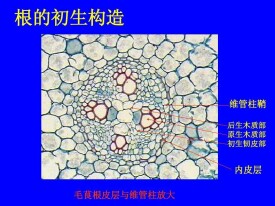
維管植物的初生生長和分化時,由原形成層分化的木質部,稱為初生木質部,其中含有纖維和薄壁組織細胞,但沒有射線,所以,不構成軸向系統和徑向系統。在莖、葉和花等部位中,初生木質部和初生韌皮部結合成束狀,即為維管束。在莖的維管束之間有薄壁組織,稱為髓射線。在根中,初生木質部在中央形成一個核心,核心中央具髓或不具髓。根據分化的早晚,初生木質部又可分為原生木質部和後生木質部兩部分。
木質部中最早形成的部分。在莖、葉中,原生木質部是在活躍伸長的組織中成熟的,因此會受到應力的影響,其中無生命的管狀分子被拉伸而毀壞。在根中,原生木質部管狀分子成熟的地方,是在伸長區以後,所以存在時間較久。原生木質部一般只有管狀分子及其包圍它們的薄壁組織。原生木質部的管狀分子,主要有環紋和螺紋加厚。
初生木質部中,原生木質部分化以後所形成的部分,如有次生木質部時,則在次生木質部發生之前產生。後生木質部一般是在初生植物體仍舊生長時開始發生,其成熟則是在伸長完成以後,因此,比原生木質部受周圍組織伸長生長的影響小。後生木質部的組成比原生木質部複雜,除了管狀分子和薄壁組織外,還有纖維。薄壁組織可以散布在管狀分子之中,也可以形成射線狀的徑向行列。在切向切面上,它們是軸向的薄壁組織細胞。後生木質部的管狀分子可以有環紋、螺紋、梯紋(圖3)以及具緣紋孔等。次生木質部維管植物次生生長時,由維管形成層產生的木質部。次生木質部分為軸向系統和徑向或射線系統。軸向系統指次生維管組織中,由紡錘狀形成層原始細胞衍生的細胞,它們的長軸與植物體軸呈平行排列。徑向系統是次生維管組織中所有射線的總稱。其細胞的長軸與體軸間成水平方向排列。
在莖或根次生木質部中的生長層。每一個生長輪是一個生長季長成的,稱為年輪。在一個生長季中,因環境條件的變化,產生一個以上的生長層,叫假年輪。在一個生長輪中,可分為春材和秋材兩部分。春材是生長層上春夏形成的木材,其細胞較大、壁薄、結構較疏鬆。秋材是生長層中秋冬時候形成的木材,比春材的細胞密而小,壁較厚,質地堅實。春材和秋材的相對數量可受環境條件的影響,也因植物種類而異。根據生長輪的數量可以推算出樹木的年齡。通過對生長輪的分析,還可了解歷史上氣候的變遷。
在種子植物木材中,軸向細胞和射線在切向切面上排列成同一水平方向,稱為疊生木材。疊生木材是由疊生形成層所產生。例如,刺槐屬的木材。非疊生木材是由非疊生形成層產生的,木材中軸向細胞和射線細胞在切向切面上並不排列成水平系列。例如胡桃屬的木材。
根和莖的木材外部,含有生活的細胞和貯存物質、並具輸導水分和無機鹽功能的部分,稱為邊材。在次生木質部內部已經停止貯藏和輸導作用的部分,稱心材,其顏色一般比邊材深。邊材的數量較為恆定,心材的相對數量在各種植物中都不同,而且還受環境的影響。一般說來,心材的數量可以不斷增加,在少數植物中,也可不形成心材。
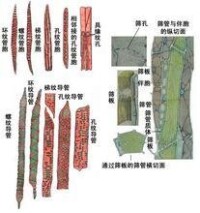
次生木質部
是沒有韌型纖維。硬材指雙子葉植物的木材。雙子葉植物的木材結構比裸子植物的變化大。原始無導管的雙子葉植物木材結構比較簡單,但在具導管的雙子葉植物木材中,還有管胞、一種或幾種纖維、軸向薄壁組織細胞和一種或幾種射線。另外,在許多雙子葉植物木材中,當導管變得不活躍以後,與導管相鄰的軸向薄壁組織細胞和射線薄壁組織細胞,可以通過紋孔形成突起,填充在導管腔內,形成侵填體。
在雙子葉植物次生木質部的橫切面上,生長輪中的導管管孔分佈相當均勻,或從早材到晚材的管孔大小逐漸變化,這種木材稱散孔材,例如毛白楊。環孔材是指雙子葉植物的次生木質部中,早材的導管管孔明顯地比晚材大,因而在橫切面上形成清楚的同心環狀排列。例如水曲柳。
應力木是莖或枝條在抵抗迫使它們傾斜或彎曲的重力作用下形成的木材。松柏類植物和雙子葉植物的應力木在產生部位與結構等方面都有相當大的差別,因此又可分為壓縮木和伸張木兩種。壓縮木,即松柏類植物的應力木。在這類植物中,傾斜或彎曲莖或枝條下側的木材與正常的木材相比,組織質地緻密,木質化程度較高,比重大,顏色深,管胞較短。在橫切面上,管胞的孔徑多為圓形。伸張木,雙子葉植物的應力木。在傾斜或向上捲曲莖或枝條上側的木材,其特點是具膠質纖維,比正常木材木質化程度低,但纖維素含量較高。伸張木也存在於根中。
陸生維管植物所需的水分,主要是由根部從土壤中吸收后,經過根和莖的木質部導管或管胞,運送到葉、花、果實等器官中。在水分的運輸途徑中,水分沿著木質部導管或管胞上升的動力,主要是蒸騰拉力。植物的蒸騰作用越強,從導管或管胞中拉水的力量也就越大,則失水越多。另外,在導管或管胞中水分之間的內聚力很大,從而形成一條連續的水柱,水柱內的內聚力可使水柱向下降。這樣上拉下拽便使水柱產生張力,水柱張力遠比水分內聚力小,因此,可使導管或管胞中的水柱不斷,這就保證了水分不斷向上運輸。儘管根壓在植物水分的正常運輸中起的作用並不大,但是多年生木本植物在早春尚未展芽之前,或土溫高,水分充足,大氣相對濕度大以及蒸騰作用小時,根壓對水分的上升就會起較明顯的作用。在各類植物的木質部中,水分運輸的速度差別較大。據統計:在木本被子植物的環孔材中,儘管孔徑較大,水流速每小時可達20~40米;散孔材中,導管孔徑一般較小,每小時只有1~6米。在裸子植物的管胞中,水的流速更慢,每小時還不到0.6米。草本植物中水流速度與環孔材樹木相近似。此外,在同一枝條上,被太陽照射部分的水流速度大於不直接照射的部分;同一植株中,晚上水流速度小於白天。水分在植物體內的運輸,主要是由下而上,除此之外,還有橫向的運輸,即通過射線和管間紋孔;以及向下運輸等途徑,但這些運輸的速度都比向上運輸慢得多。
利用木材在建築、交通、煤炭、造紙等工業方面更為重要。此外,在鋼鐵工業、農具製造業、造船業、車輛製造業、飛機製造業、膠合板、傢具、火柴、電桿、林產化工、化學纖維和其他有關生產部門,也有極廣泛的用途。
基本信息
- 界
- 植物界
- 屬性
- 管狀細胞