維管束
維管束
維管束是指維管植物(包括蕨類植物、裸子植物和被子植物)的維管組織,由木質部和韌皮部成束狀排列形成的結構。維管束多存在於莖(草本植物和木本植物幼體)、葉(葉中的維管束又稱為葉脈)等器官中。維管束相互連接構成維管系統,主要作用是為植物體輸導水分、無機鹽和有機養料等,也有支持植物體的作用。
維管束彼此交織連接,構成初生植物體來輸導水分,無機鹽及有機物質的一種輸導系統——維管系統,併兼有支持植物體的作用。
在維管束的發育過程中,初生木質部和初生韌皮部還可分為發育較早的原生木質部和原生韌皮部,以及發育較遲的後生木質部和後生韌皮部。由於早先發育的原生木質部和原生韌皮部分子,在初生植物體伸長生長時就已成熟,它們不再與周圍的細胞一起繼續伸長,因而常被擠毀,或留下原生木質部的腔隙,如玉米的莖中的維管束。
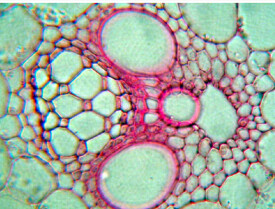
在維管束的周圍,通常由一層或數層具支持作用的厚壁組織細胞組成的維管束鞘所包圍。它們有時僅在木質部或韌皮部的一端,或同時出現在兩端。
維管植物體內木質部和韌皮部緊密結合在一起形成的束狀結構,稱維管束(vascular bundle)。 維管束是是植物進化到較高級階段出現的組織。 維管束主要由韌皮部與木質部組成,因此,維管束、木質部、韌皮部和周皮、表皮一樣等都屬於複合組織。維管束由原形成層分化而來,在不同的植物或不同的器官內,原形成層分化成的木質部和韌皮部的情況不同,就形成了不同類型的維管束。
維管束主要由韌皮部(phloem)和木質部(xylem)構成。在被子植物中,韌皮部是主由篩管、伴胞、篩胞、韌皮薄壁細胞與韌皮纖維共同構成的一種複合組織,這部分質地較韌,自上而下的主要運輸有機養料;木質部是主要由導管、管胞、木薄壁細胞與木纖維共構成的複合組織,這部分質地堅硬,自下向上的主要輸導水分,同時與礦物質的運輸、養的貯存有關。
根據維管束中木質部與韌皮部排列方式和有無形成層,從而形成以下幾種常見的維管束類型:
1.有限外韌維管束(closed collateral vascular bundle)
韌皮部在外側,木質部在內側,中間無形成層的維管束類型。常見於大多數單子葉植物和蕨類植物莖的維管束。
2.無限外韌維管束(open collateral bundle)
韌皮部在外側,木質部在內側,中間有形成層的維管束類型。如雙子葉植物和裸子植物莖的維管束。
3.雙韌維管束(bicollateral vascular bundle)
4.周韌維管束(amphicribral vascular bundle)
木質部位於中間,韌皮部圍繞於木質部四周,無形成層。常見於蕨類某些植物與百合科、禾本科、蓼科的某些植物。
5.周木維管束(amphivasal vascular bundle)
6.輻射維管束(radial vascular bundlel)
木質部與韌皮部彼此間隔交互呈輻射狀排列。常見於被子植物根的初生構造中。
維管束類型在不同植物體中存在差異,因此也是藥材鑒別的依據之一。
裸子植物和雙子葉植物維管束的木質部和韌皮部之間常有形成層,能持續進行分生生長,所以稱無限維管束或開放性維管束(open bundle)。蕨類植物和單子葉植物的維管束中不存在形成層,不能持續不斷的分生生長,故稱有限維管束或閉鎖性維管束(closed bundle)。
雙子葉植物維管束的發育過程
在維管束的發育過程中,初生韌皮部是從原形成層的外側開始,由外至內進行向心發育,先形成外方的原生韌皮部,后形成內方的後生韌皮部,其發育方式與根相同,為外始式。原生韌皮部較為柔嫩,在分化發育過程中多被破壞,因此,初生韌皮部往往完全由後生韌皮部組成。韌皮薄壁細胞散生在韌皮部中,常含澱粉、單寧、晶體等貯藏物質,韌皮纖維常成束分佈在初生韌皮部的最外側,植物種類不同,韌皮纖維的發達程度不同。
初生木質部是從原形成層的內側開始先形成原生木質部,然後進行離心發育,逐漸分化形成後生木質部,這種發育順序與根相反,為內始式(endarch)。原生木質部由環紋和螺紋導管以及木薄壁細胞組成,而後生木質部由管徑較大的網紋、孔紋、梯紋導管和木薄壁細胞組成,有時還有木纖維。
在原形成層分化形成初生維管束的過程中,初生韌皮部和初生木質部之間保留了層分生組織細胞,稱為束中形成層(fascicular cambium),因此,雙子葉植物莖中的維管束多為無限維管束,這是雙子葉植物莖進行次生生長的基礎。
單子葉植物維管束的發育過程
維管束
維管束分佈於基本組織中,數目較多,為外韌維管束。維管束的排列有兩種方式:具中空莖的禾本科植物(如小麥、水稻)中,維管束成內、外兩輪排列,外輪的維管束較小,位於莖的邊緣,大部分埋藏於機械組織中,內輪的較大,周圍為基本組織所包圍;實心結構莖的禾本科植物(如玉米、甘蔗、高粱)中,維管束星散分佈於基本組織中,外圍維管束體積較小,分佈密集,越近中央,維管束體積越大,分佈越稀疏。不論何種類型的莖,其維管束均由維管束鞘、初生木質部和初生韌皮部三部分組成。
(1)維管束鞘維管束的外面,通常由1-2層厚壁細胞組成的維管束鞘包圍(如小麥、水稻),也有的植物維管束鞘是由多層細胞組成的(如甘蔗)。
(2)初生木質部位於維管束的近軸部分,橫切面的輪廓呈“V”形。V形的基部,也即維管束向心的一面,為較早形成的原生木質部,包括1-3個環紋或螺紋導管及少量木薄壁組織,生長過程中這些導管常遭破壞,四周的薄壁細胞互相分離,形成氣腔;在V形的兩臂上,也即維管束離心的一面,各有一個後生的大型孔紋導管,以及在這兩個導管之間分佈的薄壁或厚壁細胞,有時也有管胞,共同組成了後生木質部。因此,木質部的分化是由內而外的內始式發育。
(3)初生韌皮部位於初生木質部的外方,靠近莖周方向,莖發育的後期,外側的原生韌皮部常被擠毀或殘留,內側的後生韌皮部由篩管、伴胞組成。在橫切面上,篩管較大,呈多邊形,伴胞位於篩管的旁邊,為三角形或長方形的小細胞。韌皮部的分化是由外而內的外始式發育。在初生木質部和初生韌皮部之間沒有束中形成層,為有限維管束。
維管束(vascular bundle)是維管植物(包括蕨類植物、裸子植物和被子植物)的輸導系統,在植物體內常呈束狀結構,貫穿在植物體的各種器官內,除了具有輸導功能外,同時植物器官起著支持作用。
維管組織
由木質部和韌皮部組成的輸導水分和營養物質,並有一定支持功能的植物組織。
在有次生生長的植物(大多數裸子植物和木本雙子葉植物),維管組織包括來源於原形成層的初生木質部和初生韌皮部(合稱初生維管組織)及來源於維管形成層的次生木質部和次生韌皮部(合稱次生維管組織)。在只有初生生長的植物(大多數蕨類植物和單子葉植物)維管組織只包括來源於原形成層的初生木質部和初生韌皮部。
植物系統學中,把體內具有維管組織的植物稱為維管植物。在植物進化過程中,維管組織的分化和出現,對於植物適應陸生環境具有重大意義。
維管系統
維管束彼此交織連接,構成初生植物體輸導水分、無機鹽及有機物質的一種輸導系統維管系統,併兼有支持植物體的作用。
維管束植物是植物的一個類群。蕨類植物、裸子植物、被子植物的莖葉等部位都具有維管束構造,統稱為維管束植物。
根莖葉的分化
維管束植物為多細胞植物體,細胞分化明顯,組成各種不同功能的組織。有根、莖、葉的分化。
維管束作用
植物體有由特化的細胞所組成的維管束。承擔支持和運輸的功能。
生殖器官特徵
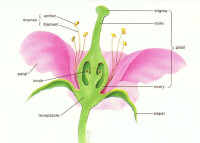
生殖器官構造複雜,特別是被子植物有完善的花。胚由受精卵細胞形成。
世代交替明顯而有規律,不受外界條件的影響而變化。無性世代佔優勢。越高等越有性生殖越退化。
維管束由初生韌皮部、束中形成層和初生木質部所組成。
初生木質部
其中初生木質部包括管狀分子(導管分子或管胞),薄壁組織細胞和纖維。
初生韌皮部
變褐色的維管束
原生木質部和原生韌皮部
在維管束的發育過程中,初生木質部和初生韌皮部還可分為發育較早的原生木質部和原生韌皮部,以及發育較遲的後生木質部和後生韌皮部。由於早先發育的原生木質部和原生韌皮部分子,在初生植物體伸長生長時就已成熟,它們不再與周圍的細胞一起繼續伸長,因而常被擠毀,或留下原生木質部的腔隙,如玉米的莖中的維管束。在維管束的周圍,通常由一層或數層具支持作用的厚壁組織細胞組成的維管束鞘所包圍。它們有時僅在木質部或韌皮部的一端,或同時出現在兩端。多年生木本植物維管束排列成桶狀。
其中初生木質部包括管狀分子(導管分子或管胞)、薄壁組織細胞和纖維。
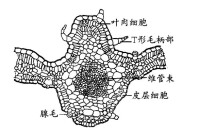
維管束
維管束彼此交織連接,構成初生植物體輸導水分、無機鹽及有機物質的一種輸導系統——維管系統,併兼有支持植物體的作用。

維管束
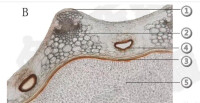
維管束
葉植物中。位於後生木質部與後生韌皮部之間的原形成層,如能繼續發育為維管形成層(即束中形成層),並與維管束之間薄壁組織產生的形成層(束間形成層)相接,向外產生次生韌皮部,向內分化次生木質部,這種能繼續生長的維管束,稱為無限維管束,常見於裸子植物和木本雙子葉植物中。
在初生植物體內,維管束相互連接、錯綜複雜。莖中的維管束進入葉子里,是通過莖皮層到葉柄基部的一段維管束,即葉跡。同樣,側芽發生后,由枝跡將莖的維管束與側枝維管束相互連接。根中的維管組織的排列與莖不同,它們之間的聯繫是通過一個過渡區,即由根的木質部束與韌皮部束交替排列的方式,逐漸轉變成莖中木質部與韌皮部內外排列成維管束的形式。總之,根通過過渡區與莖維管束相連,莖再通過枝跡與葉跡同側枝與葉子的維管束連接,這樣,在初生植物體中構成一個完整的維管組織系統,主要起輸導和支持作用。
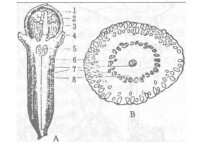
維管束
以小麥為例,小麥穎果腹部維管束 韌皮部中的篩管較多, 而處在中部的木質部中的導管較少。如花后 5 d 的穎果腹部維管束中部是 9~ 12 條導管, 其外側是呈 半圓形分佈的 4 0 條 左右的篩分子, 腹部維管束與胚囊中的胚乳細胞間有合點, 合點上方為珠心突起( 圖v1) , 該部位細胞和細胞核較大, 細胞間隙大, 有利於養分質外體輸送。小麥穎果花后 5 d 腹部維管束還沒有完全分化成型, 只有大量的果皮薄壁細胞包圍在原維管束的周圍。花后 12 d , 較完善的
輸導組織已發育形成, 篩管分子分佈於維管束外側, 左右兩部分對稱, 呈半圓形。在韌皮部薄壁細胞上方為木質部, 有 1 0 多個導管( 圖v2) 。花后16 d, 腹部維管束髮育成熟( 圖v3 ) , 具有較強的輸送養分的能力。花后 19 d 觀察穎果腹部維管束的超微結構( 圖v4) , 發現, 在穎果維管束中篩管周圍有伴胞, 形成典型的篩分子 - 伴胞複合結構, 伴胞細胞基質濃厚, 篩分子壁向內加厚, 中間細胞的液泡化更加明顯, 中央大液泡幾乎佔據細胞大部分體積, 原生質被擠到細胞的周緣。但穎果的伴胞或薄壁細胞的體積比篩管分子大, 這與莖中相反, 而且這些伴胞或薄壁細胞的細胞質濃, 線粒體密度大, 呼吸旺盛, 代謝活躍, 在功能上與莖中的伴胞不同。在其周圍的薄壁細胞原生質染色較伴胞淺, 其中有球狀的線粒體, 少量球形的質體。花后 2 8 d 穎果腹部維管束薄壁細胞外形不規則, 許多細胞壁向旁邊的細胞內突出或細胞壁內陷( 圖v5 箭頭處) , 大部分韌皮薄壁細胞胞核解體, 並逐漸消失, 線粒體數目變少說明薄壁細胞物質轉運能力開始下降。花后36 d , 輸導組織被四周的胚乳細胞擠壓, 嚴重變形和破碎, 輸導功能已基本喪失。( 圖v6) 。
基本信息
- 中文名
- 維管束
- 外文名
- vascular bundle
- 術語類別
- 有限維管束和無限維管束
- 出處
- 頂端分生組織分化的原形成層
- 功能
- 輸導功能、支持作用
- 概念
- 由韌皮部和木質部組成的束狀結構