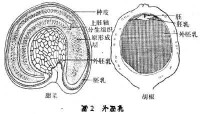
胚乳
滋養組織
胚乳(endosperm)一般是指被子植物在雙受精過程中精子與極核融合后形成的滋養組織,也稱內胚乳。這種組織既不是配子體,也不是孢子體,其染色體倍性一般為三倍體;為許多植物(如禾本科植物)種子的重要組成部分。
在裸子植物的種子中,直接由雌配子體發育而來的營養組織;以及在被子植物的種子中,通過精子與極核融合,發育而來的營養組織,統稱為胚乳。胚乳是種子貯藏營養物質的部分,為胚發育所需的營養物質,有的胚乳在胚的發育過程中,被消耗吸盡,因此。並不是所有種子都有胚乳。
胚乳是種子植物的特有結構,但被子植物和裸子植物的胚乳來源是不同的。被子植物的胚乳是受精的產物,屬於孢子體組織;而裸子植物胚乳是減數分裂的產物,屬於配子體組織。長期以來對被子植物胚乳的起源還不清楚。在雙受精中,來自同一花粉管中的兩個精細胞之一與卵細胞融合形成合子,另一個與中央細胞的次生核融合形成初生胚乳核。儘管雙受精的細胞具有相似的遺傳性並且所處環境相同,但是它們沿著兩個顯然不同的發育途徑形成了胚胎和營養胚乳。人們發現雙受精之後提出了兩個關於胚乳進化起源的假說:一種假說認為,胚乳也是一種胚胎髮生類型,但起一種利他作用;另一種假說認為胚乳是被第二次受精啟動的雌配子體的統一整體。基於現存的被子植物雙受精的發生,Friedman已經提出證據支持第一個假說。根據這個觀點,胚乳起於雙倍的額外胚胎,沿著被子植物的分化,這個雙倍的額外胚胎形成了胚胎營養組——胚乳。
根據種子裡面有無胚乳的情況。分為有胚乳種子及無胚乳種子兩類。在無胚乳的種子中,在種子形成的早期,胚乳中的營養物質被胚吸收轉移到子葉里貯藏起來,因此種子成熟后胚乳消失,子葉特別肥厚,如玉豆種子,由於胚在發育過程中,將種子的胚乳吸盡,所以種子內不存在胚乳(由子葉起著胚乳的作用).如果種子萌發時,將兩片肥大的子葉摘除后,幼芽因得不到營養物質的供給,很快就會死亡。有胚乳的種子,胚乳發達,胚較小,種子的營養物質大部分貯藏在胚乳中。在蔬菜作物中,這類種子較少,常見的稻穀、小麥等糧食作物,就是有胚乳的種子,它提供我們吃用的澱粉就是貯藏在胚乳里的。
胚乳
被子植物的胚乳是由一個精細胞與中央細胞的兩個極核或次生核受精后形成的初生胚乳核發育而成的,具有三倍染色體。
初生胚乳核的分裂早於合子的分裂,胚乳的發育總是早於胚的發育。胚乳的發育類型如下:
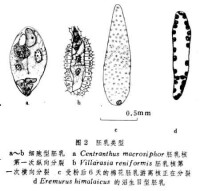
胚乳的早期發育有一遊離核時期。遊離核分裂的次數則隨植物種類而異。如咖啡屬早在4核階段就形成壁。馬利筋屬、大花草屬和還陽參屬,在8核或16核階段形成胞壁。報春花屬、錦葵屬、杧果屬、胡桃屬、蘋果屬和柑橘屬等,沿著胚囊壁可以看到幾百個胚乳核。(圖2c)
胚乳
不存在遊離核時期,初生胚乳核及其後繼的細胞分裂,有規則地形成細胞壁。根據第一次分裂形成的細胞方向,可分為縱向分裂(圖2a)和橫向分裂(圖2b)兩個亞型。五福花屬的初生胚乳細胞頭兩次分裂都是縱向的,結果形成4個柱狀細胞,到第3次分裂才是橫向的。大多數植物的初生胚乳細胞,第一次是橫向分裂。肉豆蔻科、馬兜鈴科、瓶爾草科、龍膽科和紫草科等植物,第2~3次分裂仍是橫向分裂,結果形成4個或更多的細胞系列。
初生胚乳核移到合點端後分裂,形成一個合點端小室和一個珠孔端大室。珠孔端核再行遊離核分裂,後期才形成細胞壁;合點端的核不分裂,或只行幾次遊離核分裂(圖2d)。
胚乳在被子植物種子中是普遍存在的。它的主要功能是為發育中的胚提供營養。在蘭科植物的種子中,胚乳只有很少幾個細胞。柳葉菜科、河苔草科和菱科植物的種子不具胚乳。蠶豆、豌豆和菜豆等豆科植物,種子發育初期雖有胚乳,但後期完全被發育中的胚吸收消耗了,因此種子成熟時不具胚乳或者只在種皮下殘存1~2層胚乳細胞;這種無胚乳的種子,胚具有肥厚而肉質的子葉,在種子萌發時為胚提供營養。
穀類植物胚乳最外面一層或幾層細胞,在形態與生理上高度特化,形成糊粉層。這些細胞富含糊粉粒,還含有一些圓球體。糊粉粒的主要成分是蛋白質、植酸鈣鎂、磷酯和碳水化合物。在成熟穀粒中,糊粉細胞是活化的,當籽粒萌發時,胚乳細胞的貯藏物質被糊粉層細胞分泌的水解酶所分解。
有些成熟胚乳表面,呈不同程度的凹凸不平,稱為嚼爛狀胚乳。在被子植物中約有32科植物具嚼爛狀胚乳。
胚乳吸器:最初在銀樺胚乳合點端發現的蠕蟲狀附屬物(圖3a)。以後在葫蘆科、豆科和山龍眼科等許多植物中,也相繼發現了各種各樣的胚乳吸器。Iodina rhombifolia的胚乳吸器形態複雜,甚至成珊瑚狀;Impatiensroylei的胚乳吸器呈菌絲狀分枝,並侵入珠柄組織(圖3b);Nemophila aurita的胚乳組織呈球狀,其珠孔端和合點端兩極均具胚乳吸器。在細胞型胚乳中胚乳吸器特別發達。按胚乳吸器分佈位置可將其分為合點端吸器和珠孔端吸器兩種類型。在Lomatia中,除合點端的主要吸器外,整個胚乳表面有許多單細胞的指狀突起,這就大大增加了胚乳的吸收面積。
多數被子植物胚乳,開始時是三倍體細胞。但由於胚囊發育類型不同,胚囊中極核的數目也不同,所以初生胚乳核的倍性也不相同;月見草型為二倍體;椒草型為九倍體;蓼、蔥、五福花以及德魯撒型為三倍體;皮耐亞和白花丹型為五倍體;貝母型和小白花丹型也是五倍體。即使一般為三倍體的胚乳組織,在發育過程中,也能發生倍性的改變。不少胚乳細胞有很高的倍性。例如Thesium alpinum胚乳吸器的核為384倍體,而Melampyrumpratense合點吸器的核倍性達1536倍。倍性的增加與核內有絲分裂和核融合現象有關。胚乳細胞的大小與數目也有很大變化。蔥蓮屬胚乳有特大的細胞核,核仁數目多達23個。
被子植物的胚和胚乳都是雙受精的產物,但是胚乳核和合子核的分裂並不同步,一般說來,前者分裂比後者早,所以,初期胚的發育和分化就可靠胚乳提供營養。無胚乳種子的胚可以從外胚乳甚至珠心或珠被絨氈層組織吸收營養。
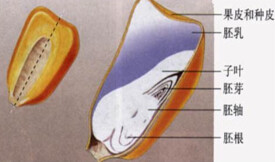
胚乳的貯藏物質主要是碳水化合物、蛋白質、脂肪和油脂。碳水化合物的基本貯藏形式為澱粉。在穀類籽粒中,澱粉是胚乳的主要貯藏物質。人們食用糧食(例如水稻、小麥和玉米等)的主要部分,也是種子的胚乳部分。在糊粉層細胞中還含有大量蛋白質。蓖麻胚乳是典型的高脂肪胚乳。在胡椒科、莧科、馬齒莧科、白花菜科、姜科和美人蕉科,營養物質也可以貯藏在外胚乳中。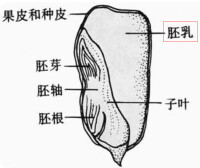
胚乳
植物胚乳的化學成分非常複雜,例如椰子乳汁,除含無機鹽、20種氨基酸和其他含氮化合物、有機酸、維生素、糖等以外,還含有一些激素。由組織培養試驗表明,椰子乳汁起生長誘導作用的主要成分是己糖醇、肌醇、二苯尿和嘌呤等,其中嘌呤成分為玉米素核糖苷,它具有高度的細胞分裂素的活性。起主要作用的氨基酸是苯丙氨酸,對細胞分裂有起動作用。發育中的玉米胚乳含有大量碳水化合物、氨基酸、蛋白質、吲哚乙酸和嘌呤等,70年代以來還發現一種細胞分裂素──玉米素。
根據G.L.戴維斯的報告,在288科被子植物中,161科為核型胚乳,72科為細胞型,只有17科為沿生目型胚乳。細胞型胚乳多集中於雙子葉植物,單子葉植物中只有天南星科和浮萍科為細胞型。同樣,在17科沼生目型胚乳中,單子葉植物佔14科。從胚乳類型之間的系統發育來看,一般認為沼生目型胚乳是中間類型。K.R.斯波恩應用統計學方法發現,核型胚乳與一系列具有原始特徵的植物關係密切,認為是由核型向細胞型過渡。B.G.L.斯沃米和P.M.迦納珀蒂根據導管穿孔類型,以是否為梯狀穿孔導管或單穿孔,來計算統計學上的相關性,發現核型胚乳與具單穿孔的植物有比較肯定的關係。他們認為導管分子起源於具梯狀梯紋紋孔的管胞;而單穿孔板比梯狀穿孔板較為特化,因此,主張由細胞型向核型過渡。到底哪一種胚乳類型較原始,至今還沒有肯定的答案。
從1933年L.蘭普和C.O.米爾利用植物組織培養方法,培養玉米幼嫩胚乳起,到1979年才有胚乳植株產生,到20世紀80年代只有少數胚乳植株培養成功,例如Putranjiva roxburghii、水稻、蘋果、柚、檀香、大麥、馬鈴薯和獼猴桃等。胚乳植株不一定是三倍體植株,而往往是混倍體。由於染色體數目和形態發生變異,胚乳的試管培養可望得到新類型的植株。胚乳植株的培養成功,說明胚乳細胞與二倍體或單倍體細胞一樣具有全能性(見潛能)。
基本信息
- 中文名
- 胚乳
- 外文名
- endosperm
- 別名
- 內胚乳
- 性質
- 精子與極核融合后形成的滋養組織