共找到2條詞條名為磷脂醯肌醇的結果 展開
- 磷脂醯肌醇
- 肌醇磷脂
磷脂醯肌醇
磷脂醯肌醇
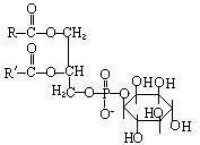
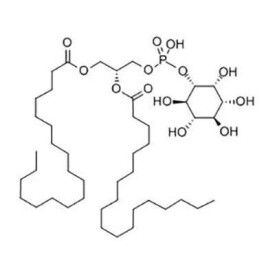
英文名稱:Phosphatidylinositol,簡稱: PI。
徠PI主要由兩部分組成的,一是磷酸1,2-二脂醯甘油,二是肌醇(inositol)。它在細胞中對於細胞形態、代謝調控、信號傳導和細胞的各種生理功能起著非常重要的作用。
英文名稱:Phosphatidylinositol
化學途徑
是G蛋白偶聯受體的信號轉導通路中的一種途徑,在信號通路中胞外信號分子與細胞表面G蛋白耦聯型受體結合,激活質膜上的磷脂酶C(PLC-β),使質膜上4,5-二磷酸磷脂醯肌醇(PIP2)水解成1,4,5-三磷酸肌醇(IP3)和二醯基甘油(DG)兩個第二信使,胞外信號轉換為胞內信號,這一信號系統又稱為“雙信使系統”(Double Messenger System)。
DG結合於質膜上,可活化與質膜結合的蛋白激酶C(Protein Kinase C,PKC)。PKC以非活性形式分佈於細胞溶質中,當細胞接受刺激,產生IP3,使Ca2+濃度升高,PKC便轉位到質膜內表面,被DG活化,PKC可以使蛋白質的絲氨酸/蘇氨酸殘基磷酸化使不同的細胞產生不同的反應,如細胞分泌、肌肉收縮、細胞增殖和分化等。DG的作用可用佛波醇酯(phorbol ester)模擬。
DG通過兩種途徑終止其信使作用:一是被DG-激酶磷酸化成為磷脂酸,進入磷脂醯肌醇循環;二是被DG酯酶水解成單酯醯甘油。由於DG代謝周期很短,不可能長期維持PKC活性,而細胞增殖或分化行為的變化又要求PKC長期活性所產生的效應。現發現另一種DG生成途徑,即由磷脂酶催化質膜上的磷脂醯膽鹼斷裂產生的DG,用來維持PKC的長期效應。
首先由激活的SrcPrK和ZAP-70通過LAT使膜結合的磷脂酶C(PLC)分子丁鏈上的酪氨酸殘基發生磷酸化。磷酸化的PLC—γ發揮酶活性,使底物二磷酸磷脂醯肌醇(PIP2)水解成兩個成分:三磷酸肌醇(IP3)和二醯甘油(DAG)。IP3可迅速地從膜內側向胞質溶膠中擴散,一方面打開細胞膜上的鈣通道使Ca2+進入細胞內,同時開啟細胞內鈣池(內質網)增加Ca2+—的釋放,協同提高胞內遊離鈣的濃度。胞質Ca2+含量的上升,激活一種稱為鈣調蛋白(camodulin)的Ca2+結合蛋白,後者可調節其他酶類的活性,並最終導致鈣調磷酸酶的激活。
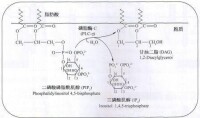
T細胞抗原激活信號轉導磷脂醯肌醇途徑
鈣調磷酸酶是一種絲、蘇氨酸磷酸酶而不是PTK。另一方面,與胞膜內側相聯的DAG則直接激活PKC。後面熔會捍到,鈣調磷酸酶和PKC主要分別活化兩種重要的轉錄因子NF—AT和NF—cB。因而在這一條信號轉導的下游通路中,實際上再一分為二,形成鈣調磷酸酶參與的途徑。和PKC介導的途徑。由於一個PLCγ分子可以產生很多的IP2和DAG,這就放大了傳人的抗原識別信號.並保證其轉導的有效性。
Ca2+活化各種Ca2+結合蛋白引起細胞反應,鈣調素(calmodulin,CaM)由單一肽鏈構成,具有四個鈣離子結合部位。結合鈣離子發生構象改變,可激活鈣調素依賴性激酶(CaM-Kinase)。細胞對Ca2+的反應取決於細胞內鈣結合蛋白和鈣調素依賴性激酶的種類。如:在哺乳類腦神經元突觸處鈣調素依賴性激酶Ⅱ十分豐富,與記憶形成有關。該蛋白髮生點突變的小鼠表現出明顯的記憶無能。
IP3信號的終止是通過去磷酸化形成IP2,或被磷酸化形成IP4。Ca2+由質膜上的Ca2+泵和Na+-Ca2+交換器將抽出細胞,或由內質網膜上的鈣泵抽進內質網。
基本信息
- 中文名
- 磷脂醯肌醇
- 外文名
- Phosphatidylinositol
- 組成
- 磷酸1,2-二脂醯甘油和肌醇
- 化學途徑
- G蛋白偶聯受體的信號轉導通路
- 分子量
- 867.1383
- CAS號
- 97281-52-2
- 分子式
- C45H87O13P
- 簡稱
- PI