疑源類
疑源類
疑源類主要是不同門類的單細胞藻。可能包括一些溝鞭藻(無溝鞭藻類的鑒別特徵)和綠藻等,絕大多數為飄浮類型。中央腔封閉或與外部相通,其對稱性、形態、結構和紋飾等變化不一,以球形體和刺狀體為主,直徑從十幾微米至數百微米不等,通常小於100微米。
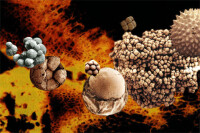
疑源類生物
對疑源類的研究開始於20世紀30年代。20世紀50年代時,隨著石油勘探的大量進行,疑源類開始被大量研究。1963年,Evitt提出疑源類這個概念(group Acritarcha,出自希臘語 akritos,不確定或混淆的;arche,起源),並將其定義為“未知並可能具有不同生物親緣關係的微體化石,由一層或多層壁包圍中央腔組成,壁的主要成分是有機物. . . . . . ”,並確定了疑源類的基本分類方法。疑源類的研究在20世紀60—70年代進入了第二階段。這時人們認識到疑源類是一類重要的化石類群,特別是在生物地層學方面,因此廣泛開展疑源類分類和系統研究,大量的疑源類被命名和描述。80年代后,對疑源類進入第三階段,對疑源類的研究被廣泛應用到系統發生學、生態學、古地理學和古環境學等方面的研究上。
疑源類的大小的變化範圍極大,大小從小於10μm到大於1mm都有,大多數種類的大小在15μm—80μm 之間。它們的外形變化也很大,圓形、橢圓形、圓盤形、長形或多角形的中空殼體或中心體,在殼體表面還可能有突起、網脊、顆粒、翼、膜等外圍修飾。有的種類外殼是多重的。
1963年時,Evitt就明確指出疑源類是一個非正式的、實用的、沒有屬以上單位的分類類目。他建議疑源類使用國際植物命名法命名形態屬和形態種,而使疑源類不被歸入任何特定的生物門類之下。雖然這種分類方案可能並不能反映分類單元中系統發生關係,但基於形態相似性的分類類群可能仍代表了自然類群。應用最廣的疑源類分類方案之一是Evitt和Sarjeant(1963) 提出的基於殼體形狀和突起形態劃分的13個亞類,用“-morphitae”作為結尾。由於亞類不是命名法規規定的術語,使得屬可以在亞類間轉移或當其親緣關係確定後轉移至林奈系統的科下。之後又對增加和減少了一些亞類。
疑源類最早可能在18億年前的古元古代出現。而可確認的疑源類則出現於15億年前的中元古代。晚元古代時,疑源類快速分化。但在新元古代末期,疑源類可能經歷了一次大滅絕。古生代早期疑源類的種類和數量不斷增長,雖然在奧陶紀末期又經歷了一次滅絕事件,但此後又快速增長。泥盆紀滅絕事件后,疑源類的種類和數量就一直處於很低的狀態。第三紀時,疑源類也處於消退狀態,唯中新世略有恢復。疑源類主要出現在海相沉積地層中,在湖相和河相地層中也有發現。

基本信息
- 中文名
- 疑源類
- 外文名
- Acritarchs
- 定義
- 有機壁、親緣關係不明的微體化石