溝鞭藻
溝鞭藻
溝鞭藻屬甲藻門(Pyrrophyta),溝鞭藻綱(Dinophyceae),現為溝鞭藻門(Dinoflagellata),為一種原始的真核有機體(Evitt,1985;Fensome,Riding & Taylor,1996)。單細胞為主,浮遊,個體微小,一般為20-250µm。溝鞭藻是構成現代海洋中食物鏈的重要基礎部分(有人稱為“海里的草”),也是主要的氧氣製造者,在淡水和半鹹水的湖泊等水體中也存在。最早的化石紀錄為上三疊世。更重要的是石油和天然氣等烴類礦產資源的生產者(郝詒純和茅紹智,1993)。個體微小,具抗酸性外壁,廣泛的地理分佈和迅速的演化——生物地層上具有重要的價值。現存溝鞭藻120多屬,1000餘種;化石溝鞭藻400餘屬,2000餘種。
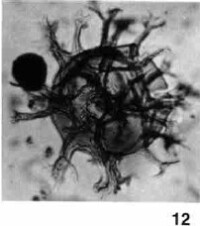
溝鞭藻
2、橫向分佈的橫鞭毛和縱向分佈的縱鞭毛,分別從橫溝、縱溝內的鞭毛孔中伸出。橫鞭似帶狀,嵌於橫溝內,環繞著細胞體,它的運動控制著細胞作螺旋式前進;縱鞭呈線狀,嵌於縱向的縱溝中。典型的從細胞的後方伸出,它的運動只能使細胞產生某種向前的運動,更確切地說,縱鞭起著舵的作用。溝鞭藻之所以能在水中作最快達1m/h的運動,就是靠著這兩根鞭毛相互配合運動的結果。
6、某些溝鞭藻還具有發光性。
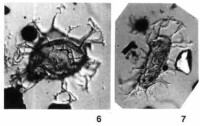
溝鞭藻
溝鞭藻
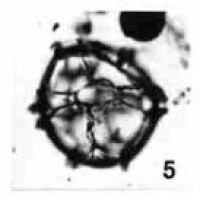
溝鞭藻
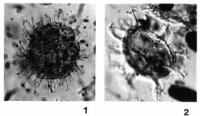
溝鞭藻
海洋是溝鞭藻生活的“大本營“,除海洋外,溝鞭藻也見於淡水湖泊、河流、污水、泥塘、陰溝或半鹹水瀉湖中,甚至在雪和海、湖沙灘的間隙水中存在(Fensome et al.,1996)。
2、營養方式
在溝鞭藻的活動期,約有一半的溝鞭藻為自養型(通過光合作用產生自己的食物)另一半則為他養型,其中多數以攝取固體生物為生(類似於動物),還有一些營共生或寄生生活。
3、控制溝鞭藻生長及分佈的因素
透光性、溫度、鹽度、營養供給、營養方式、上涌流、潮的影響、污染等。一般生活在相對較淺的水中,在正常光照條件下,18米-90米達到最大的濃度,但當水中含有大量泥沙等懸浮物時,這個深度可降到1-10米。許多溝鞭藻具有較廣的溫限性能,可忍耐的溫度為1-35℃,但最適宜在18-25℃水體中發育。有些種則要求一定的溫度條件,如暖水種和冷水種。海洋溝鞭藻發現於任何緯度的水域中,但在熱帶分異度呈現最大值,在近海溫帶水中則丰度最大。寒冷水體中,屬種單調但個體豐富。溝鞭藻類作為整體可以生活在不同的鹽度。溝鞭藻的生存需要一定的溶解養料。在發育上涌洋流的地區浮游生物繁盛,富含重要的磷和氮元素,使溝鞭藻興旺繁盛。相反,在遠離大陸、又無上涌洋流而缺乏營養成分的大洋中心,一般缺少或不存在溝鞭藻。赤潮即“開花”——某些種的高速分裂繁殖在海洋中產生的高密集溝鞭藻,可使海水呈紅褐色或牛奶狀。因為溝鞭藻含有毒素,大量的毒素可殺害貝類、魚蝦類,進一步危及人類。赤潮在熱帶水中可能會在任何時間產生,在溫帶水中只發生在夏季。
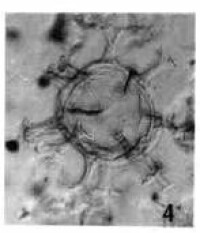
溝鞭藻
典型陸相溝鞭藻特徵
組合丰度不高;屬種分異度極低於海相,屬種的優勢度很高;以原層式近似孢囊為主,腔孢囊的內外層壁分化不明顯,刺孢囊多不發育。同一屬種陸相中的個體較小古口及板式顯示不清,古口不發育,多為裂縫狀外壁薄,有機染色不上色。組合中多見角藻型和多甲藻型孢囊,旋溝藻型少見,有也為短刺類型。
化石溝鞭藻最早由德國微生物學家Ehrenberg於1836發現。前一百年,化石溝鞭藻的研究雖有進展,但仍限於少數人的孤立工作和純學術的研究。第二次世界大戰后,由於各國對烴類能源和海洋資源的大力勘探和開發才促使化石溝鞭藻的研究迅速發展並有了新的突破,尤其是在1968年以後,隨著深海鑽探計劃和大洋鑽探計劃的實施,大大地推進了溝鞭藻生物地層的研究,進一步密切了溝鞭藻與古海洋學、海洋地質學的關係。從化石溝鞭藻的發現、分類及演化、生物地層及古生態等方面研究的進展,大致可劃分為如下四個階段:
第一階段:溝鞭藻的發現及描述階段(19世紀早期——20世紀50年代中期)
Ehrenberg(1795-1867)在1836年證明某些沉積物可以完全由微小生物的化石組成。在研究白堊紀燧石層的薄片時,他第一次描述了幾個微體化石組合,並在其中首次發現了化石溝鞭藻,命名、描述並圖示了化石溝鞭藻的某些種。並認為一些化石溝鞭藻可歸入現代屬Peridinium。出現在同一組合中的另外一類帶刺的,與現代溝鞭藻不相似的化石,他把它們歸入現代屬“Xanthidium” (一種淡水鼓藻)。他並認為“Xanthidium”與他歸入Peridinium的化石都是硅質的。1845年Mantell通過加熱使以上兩類化石變成焦炭,才證實了那些化石是由有機質組成的早期的研究對象完全是燧石和燧石層的薄片,其中有時富含微體化石,但相當部分經常不含化石,而且,所含化石在薄片中往往被太厚的岩石掩蓋或被礦物質污染。十九世紀三十年代中期,Eisenack在經鹽酸或醋酸處理過的石灰岩的不溶殘餘物中,發現了富含刺球類和其它孢粉型的有機質微體化石。第二次世界大戰後,常規的浸解(離析)技術除了除鈣的鹽酸處理外,還包括了除硅的氫氟酸處理,氧化,重液浮選和過篩。此外,光學設備的精度顯著提高。這些均促使對刺球類工作的不斷深入。無疑,溝鞭藻的研究也越來越引起關注。
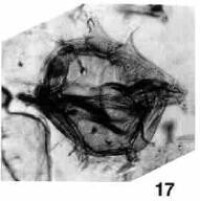
溝鞭藻
Deflandre和Cookson(1955)發表的一篇研究澳大利亞中生代晚期及第三紀組合的論文,標誌了溝鞭藻研究在南半球的開始。Eisenack和他的三位學生撰寫了一系列關於德國北部中生代和第三紀溝鞭藻組合的論文。另一位德國人Gerhard von der Brelie(1958)的研究指出,溝鞭藻在繪製第三紀海岸線時可以用來作為海相的指示者。他的工作標誌了溝鞭藻對於古環境應用的開始。在英國 Downie(1957)研究了英國南部上侏羅統Kimmeridge階頁岩中的化石溝鞭藻和刺球類,並創建了專門研究化石溝鞭藻的學校。溝鞭藻在地層劃分、對比上的意義逐漸被認識並得到應用。Wall和Dale從現代海底沉積和海水中獲得了溝鞭藻孢囊,對其進行了系統的研究,成功地使溝鞭藻孢囊發生脫囊作用(excystment)。通過實驗證明Spiriferites ramosus 的孢囊脫囊后易形成Gongaulax spinifera 的動細胞,從而決定性地證實了大多數中生代和新生代的刺球類是溝鞭藻的孢囊。該階段溝鞭藻的化石記錄得以廣泛報道。著名的出版物包括Sarjeant關於三疊紀的;Dodekova, Gocht和Gorka關於侏羅紀的;Clarke 和 Verdier ,Cookson 和Hughes及Millioud關於白堊紀的及Drugg關於新生代的。
第三階段:化石溝鞭藻的編錄及生物地層的廣泛應用階段(1967—1977)
溝鞭藻廣泛應用於地層劃分和對比,在歐洲和北美等地相繼建立了和有孔蟲、鈣質超微化石或菊石帶可對比的有關地質時代的溝鞭藻帶/組合。詳細的生物帶研究始於Clarke和Verdier(1967)研究英國南部The Isle of Wight上白堊統溝鞭藻時建立的5個溝鞭藻化石帶。Johnson 和Hill(1973)在加拿大北極地區建立了侏羅紀的5個溝鞭藻化石帶;Caro(1973)在研究西班牙北部Campo剖面時,建立了古近紀5個溝鞭藻化石帶。由於研究程度的提高以及研究範圍的擴大,新屬和新種的數量大大增加。1954年僅52屬209種,到八十年代末增至414屬2210種,大量溝鞭藻研究成果出版問世。Downie和 Sarjeant、Evitt、G. Williams,E. Kidson和 Sarjeant 發表了關於系統描述形態術語的專著。Eisenaek 和 Kjellström 出版了圖文並舉的溝鞭藻新屬種編錄(Katalog)。Lentin 和Williams編輯、出版了溝鞭藻化石種的索引(Index)。Sarjeant和 Downie提出了新的屬級以上的分類方案。Evitt每年在他的斯丹佛大學舉辦溝鞭藻短訓班,訓練、培養了大量從事溝鞭藻研究的專門人才。
第四階段:溝鞭藻的綜合性研究階段(1977—現在)
中、新生代溝鞭藻化石的生物地層格架已逐步建立起來,並可與菊石、有孔蟲和超微化石的生物地層格架相對比。研究技術的提高進而加深了對溝鞭藻形態學和系統發生學研究的程度。溝鞭藻的古生態研究進入了廣泛的探索,取得了卓有成效的成果。溝鞭藻在古生物地理、古環境、古氣候等方面的應用都進行了開創性的嘗試。
一、生物地層學方面
Williams和Bujak在1977年首次建立赤道及中緯度地區的中、新生代的溝鞭藻化石帶之後,於1985年根據許多重要屬種的時代延續提出各中、新生代的“全球溝鞭藻組合帶”。對不同緯度分佈的有關屬種的地史分佈做了總結(Williams等,1993,1999,2001)。這些為中、新生代生物地層格架的確立打下了堅實的基礎。溝鞭藻屬種的編錄(Katalog)和索引(Index)仍繼續進行(Fensome,Gocht et al.,1991,1993,1995,1996;Lentin & Williams,1977,1981,1985,1989,1993,1998),大大促進了各國學者間的交流和溝鞭藻研究的全球化。二、譜系、演化及與分類
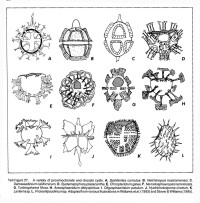
某些溝鞭藻類別的譜系和演化方面的研究有了進展,如Eatonicysta-Areosphaeridium譜系和Eneigynia-Wanaen譜系的提出(Eaton,1971;Fensome,1981)。對溝鞭藻的演化趨勢也提出了不同的模式,如板片增加或板片減少模式等(Loeblich,1976;Dorhofer & Davies,1980;Eaton,1980)。對屬種描述進行了系統總結,且附註了每個屬種的素描(Stover & Evitt,1978;Eaton,1980;Fensome 等,1993)。 Norris(1978a,1978b)提出了一個綜合的分類方案,接近於一個系統發生的分類方案,拒絕接受對孢囊和動甲使用二元分類術語。根據板式、總體形狀及角的發育、收縮程度、外壁分層性、古口類型及紋飾分佈特徵,對三疊紀至第四紀具有有機質壁的孢囊提出了一個較為完整的屬以上的分類方案。Fensome、Taylor等(1993)更加強調把化石溝鞭藻和現生的溝鞭藻結合起來,注重壁的結構,特別是板式和擬板式。主要依據核的特徵把溝鞭藻動物門分為兩個亞門和四個綱,根據總體上板式類型分為五個亞綱,又主要依據板式模式的明顯不同分為十四個目、十個亞目、五十六個科,根據板式的明顯不同以及細胞或孢囊形態的明顯不同劃分出22亞科。
三、生物地理分區
主要開展了中生代、第四紀及現代的生物地理分區,即對不同地區、不同時代某些類型及組合的地理分佈進行了研究(Fisher & Riley, 1980;Davies & Norris, 1980;Lentin & Bujak,1980;Goodman,1988;Dorhofer,1980;Mao & Mohr,1992;Wall et al.,1977;Williams & Bujak,1977;Harland,1983,1991,茅紹智,2000),並對控制地理分佈的可能的原因從環境的角度作了探討。
四、溝鞭藻古生態與古環境研究
把化石溝鞭藻作為海相的標誌,但隨著時間的推移,化石溝鞭藻在非海相的沉積中產出不斷被證實。第一個可靠無疑的紀錄是來自美國佛蒙特州的漸新統(A.Treverse,1955),在波蘭早中新世陸相地層和沼澤沉積中也有報道(Macko,1957);澳大利亞西南的湖泊和三角洲沉積(1960)美國第四紀湖泊沉積(1965)中發現淡水化石溝鞭藻;我國黑龍江、內蒙、湖北及廣東等地陸相白堊紀地層中也發現了溝鞭藻化石(茅紹智等,1990, 1999)。海相和陸相溝鞭藻組合的面貌是存在差異的,非海相的半鹹水和過渡相的溝鞭藻組合體現了典型海、陸相組合之間的過渡面貌。對溝鞭藻的古生態的研究始於三十多年前(Gocht, 1969),目前仍處於深化探索階段。對現代海洋中的溝鞭藻研究證明,水體鹽度、水溫和水深或離岸的距離等環境因素影響著居群的組成,溝鞭藻似乎較其它微體化石更能忍受鹽度的變化。Wall等(1977)根據大西洋及附近地區現代沉積物中溝鞭藻的特點,利用聚類分析方法,分析了不同緯度區的溝鞭藻組合面貌以及不同孢囊屬種的生態環境特點,提出了緯度與海岸線的距離遠近是控制孢囊組合面貌的兩個重要因素。Marret(1994)對現代肯亞海灣的沉積物中溝鞭藻的研究表明,溝鞭藻的丰度受海水深度和海水表面鹽度影響,而種的分佈則是由水體中的營養水平所控制。Oboh-Ikuenobe等(1999)認為營養水平在控制溝鞭藻的丰度和類型上比鹽度和水深更重要。溝鞭藻的絕對丰度、分異度及組合變化可以反映環境的變化。Wall 等(1977)、Lewis 等(1990)、Powell 等(1992)、Dale和Fjellsa(1994)、Dale(1996)討論了現代及第四紀上升流和古生產力方面溝鞭藻孢囊的潛在的作用。利用溝鞭藻研究近期的各種環境變化(富營養、工業污染、短期氣候變化),擴大了溝鞭藻的應用(Thorsen & Dale,1997;Firth & Clark,1998)Köthe(1990)根據主要的生態因素在德國西北部建立了一個溝鞭藻的古生態模式。Stover等(1994)在研究比利時西北部早漸新世Boom Clay Formation的溝鞭藻,探索其對層序地層的應用時,採用Köthe的模式,並作了適當修改。 Edwards(1992)根據大量的資料,分析了若干溝鞭藻屬種最適宜的環境條件(包括水深、冬、夏季表層水溫等)。Brinkhuis(1994)根據形態上與現存溝鞭藻類似或具有現代溝鞭藻的對應種的溝鞭藻孢囊分佈的定量研究得出了漸新世溝鞭藻的分佈模式。
五、古氣候研究
Wall等(1977)、Harland(1983)和Dale(1996)等溝鞭藻學家研究了現代溝鞭藻的分佈與溫度的關係。Dale(1996)依據現代溝鞭藻孢囊的分佈建立了大西洋從高緯度到低緯度不同地區的溝鞭藻孢囊的冷暖不同的主要組合特徵,並討論的一些對溫度反映敏感的種在不同緯度的分佈情況。Brinkhuis和Biffi(1993)根據溝鞭藻低緯度分子相對丰度的變化和明顯入侵地中海地區的許多中高緯度的分子相對比,在始新世與漸新世過渡帶內識別出兩個冷暖旋迴。Brinkhuis等(1998)依據溝鞭藻組合的變化重建了白堊紀與第三紀界限附近海水表面溫度。
1、《微體古生物學教程》郝詒純 茅紹智 中國地質大學出版社
2、南海ODP184航次1148站位漸新世溝鞭藻生物地層 李 傑 茅紹智
