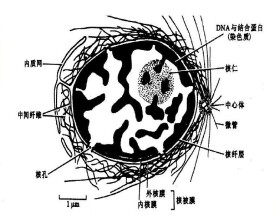
核仁
核仁
徠核仁(nucleolus)通常表現為單一或多個勻質的球形小體,是真核細胞間期核中最顯著的結構。核仁的大小、形狀和數目隨生物的種類、細胞類型和細胞代謝狀態而變化。蛋白質合成旺盛、活躍生長的細胞如分泌細胞、卵母細胞,核仁大,可佔總核體積的25%,不具蛋白質合成能力的細胞如肌肉細胞、休眠的植物細胞,核仁很小。
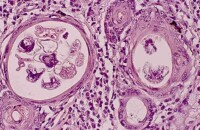
核仁內部
在光鏡下的染過色的細胞內,或者相差顯微鏡下的活細胞中,或者分離細胞的細胞核內,都容易看到核仁,它通常是單一的或者多個勻質的球形小體,呈中圓形或橢圓形的顆粒狀結構,沒有外膜。
核仁超微結構有纖維中心(FC)、緻密纖維組分(DFC)、顆粒組分(GC)三個特徵性的區域。
u纖維中心(FC):被DFC包圍的一個或幾個低電子密度的圓形結構區域,主要成分為rDNA,可看成rRNA.基因儲存的場所。
u緻密纖維組分(DFC):由緻密的纖維構成,是核仁中電子密度最高的部分,是新合成的rRNA.及其結合蛋白存在的場所,rRNA.剪切和加工場所。
u顆粒組分(GC):由核糖核蛋白顆粒構成,是正在加工成熟的核糖體亞單位的前體顆粒,容易被蛋白酶和RNase(核糖核酸酶)。
核仁組成成分包括rRNA,rDNA和核糖核蛋白。核仁是rRNA基因存儲,rRNA合成加工以及核糖體亞單位的裝配場所。
核仁的大小、形狀隨生物的種類、細胞類型和細胞代謝狀態而變化。蛋白質合成旺盛、活躍生長的細胞,如分泌細胞、卵母細胞的核仁大,可佔總核體積的25%不具蛋白質合成能力的細胞,如肌肉細胞、休眠的植物細胞,其核仁很小。
各種生物的核仁數目一般是一定的,如非洲爪蛙有兩個核仁。人類的細胞只有一個核仁。
在細胞周期過程中,核仁是一個高度動態的結構,在有絲分裂期間表現出周期性的消失與重建。細胞分裂時,核仁消失,分裂結束后,兩個子細胞分別產生新的核仁。核仁是一種動態結構,隨細胞周期的變化而變化,即形成——消失——形成,這種變化稱為核仁周期。在細胞周期中,核仁進行分離和重新聚合的過程。當細胞進入有絲分裂時,核仁首先變形和變小;其後染色質凝集和停止核糖核酸(RNA)合成,包含有核糖體RNA(rRNA)基因的DNA袢環逐漸收縮回到相應染色體的核仁組織區;核膜破裂進入中期,這時核仁消失;在有絲分裂末期時,核仁組織區DNA解凝集,rRNA合成重新開始,極小的核仁重新出現在染色體核仁組織區附近。核仁的重建過程與原有的核仁組分的協助和參與有關。核仁形成的分子機理尚不清楚,但需要rRNA基因的激活。
核仁組織區(nucleolus organizer region):即rRNA序列區,它與細胞間期核仁形成有關,構成核仁的某一個或幾個特定染色體片斷。這一片段的DNA轉錄為rRNA。
人類的核仁組織區位於10個(5對)染色體的一端,新生的核仁本來有10個,但都很小,很快融合成一個大核仁。
概況
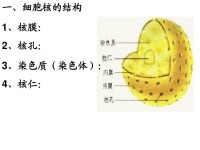
核仁
RNA基因的轉錄
RNA基因定位在核仁組織區,該區域的基因編碼18S、5.8S和28S RNA,這三個基因組成一個轉錄單位。rRNA基因在染色質軸絲上呈串聯重複排列;沿轉錄方向,新生RNA鏈從DNA長軸兩側垂直伸展出來,而且從一端到另一端有規律地逐漸增長,形成箭頭狀,外形似“聖誕樹”。每個箭頭狀結構代表是RNA基因轉錄單位,在箭頭狀結構間存在著裸露的不被轉錄的DNA間隔片段。
RNA前體的加工成熟
DNA轉錄單位轉錄出45S RNA前體,很快前體被甲基化,並剪接為41S RNA前體;41S RNA在相同的剪接位點可按照不同的剪接順序產生不同的中間前體RNA,最終將41S RNA前體剪接為28S、18S和5.8S RNA。
核糖體亞單位的組裝
核仁的主要功能之一就是組裝蛋白質合成的機器——核糖體。核糖體的生物發生包括RNA的合成、加工和核糖體亞單位的裝配等過程,是一個向量過程。從核仁纖維組分開始,再向顆粒組分延續。核糖體小亞單位成熟較早,大亞單位成熟較晚。核糖體的成熟作用僅發生在它們的亞單位被轉移到細胞質以後,兩個亞單位只有分別通過核孔進入細胞質中,才能形成功能單位。這可阻止有功能的核糖體與核內加工不完全的hnRNA分子接近。大亞單位中的5S RNA基因並不定位於核仁上,不同物種其位置不同。
真核細胞細胞核內含核糖核酸(RNA)的結構,參與核糖體核糖核酸(rRNA)的合成和核糖體的形成。在光學顯微鏡下觀察,核仁一般為圓球形或卵球形的嗜鹼性結構,常處於細胞核內偏中心的位置。其大小因細胞類型和生理狀態不同而有很大差別。在蛋白質合成旺盛的細胞,通常核仁大,反之則體積小;前者如分泌細胞、神經元和腫瘤細胞等,後者如分裂球和肌細胞等。由於核糖體是細胞內蛋白質合成的場所,而核仁的功能又是合成rRNA,因此核仁常被稱為“核糖體工廠”。核仁所含RNA約佔其乾重的5~10%,與蛋白質相結合形成核糖核蛋白(RNP)。此外還含有少量DNA,主要存在於核仁相隨染色質部分。
很多類型細胞的核仁是由核仁絲組成的網織狀結構,其中空隙填充著無定型基質。在電子顯微鏡下觀察,核仁絲是由緊密交織的、5納米的纖絲所組成,其間散在著15~20納米直徑的緻密顆粒。核仁區周緣的一部分圍繞著核仁相隨染色質(圖1)。核仁相隨染色質是異染色質,但其中一部分以襻的形式伸展到核仁內部的是常染色質。以上三種成分所組成的三維結構,即使在同一個細胞中也會因其生理狀態不同而發生改變。許多證據表明,核仁纖絲含有顆粒成分的前體。例如,用放射自顯術結合生化分析的結果,表明為rRNA編碼的基因(rDNA)的最初產物(45S RNA)首先出現於纖絲成分,隨後在核仁內被加工,然後才能定位在顆粒成分。用電子顯微鏡三維重建技術發現許多細胞的核仁與核膜存在直接或間接的連接。間接連接包括與核膜的多種衍生結構,如環形片層、核芽、核通道和核內小管等的連接。上述現象特別易見於生長旺盛的細胞,因此可能有利於加速細胞質與核仁物質的交換。
在細胞有絲分裂過程中,核仁出現周期性變化。一般在分裂前期逐漸消失,其纖絲和顆粒成分散失於核質之中;在分裂末期又重新出現。核仁的形成常與特定染色體的一定區域密切相關。在核仁消失后,這一區域就形成染色體上的次縊痕。次縊痕一般起著核仁組織區的作用。許多動、植物細胞只有一對這樣的染色體,稱為隨體染色體。但也有不少種類的細胞有幾對,甚至許多對這樣的染色體參與核仁形成,例如,人類細胞的第13、14、15、21和22對染色體上就都存在核仁組織區。實際上,核仁可被視為這些染色體的延伸,常表現很強的融合傾向,因此在許多有多對核仁染色體的細胞核里,常常只形成一個核仁。例如人淋巴母細胞在分裂前期,幾條核仁染色體都以其核仁組織區附著於同一個即將消失的核仁上。
核仁組織區存在rRNA的基因rDNA;在所有真核生物的細胞中,這種基因約有50~1000個相同拷貝,因而得以維持適當數量的rRNA的合成。真核細胞rDNA的每一個單位除含有轉錄區外,同時還有許多非轉錄區,稱為間隔順序(圖2),不同種屬、或同一種屬不同個體,甚至同一個體細胞的不同rRNA單位之間,這種間隔順序的長度往往差別很大。這種差別的意義及間隔順序的功能。
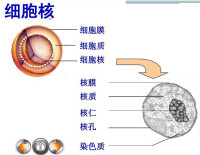
核仁
核糖體是細胞內合成蛋白質的工廠,在一個旺盛生長的細菌中,大約有20000個核糖體,其蛋白占細胞總蛋白的10%,RNA占細胞總RNA的80%。
核糖體的結構
核糖體含40%的蛋白質、60%的RNA,蛋白按照一定的順序與RNA結合,組成兩個核糖體亞單體,其中RNAs是骨架結構,有些蛋白質不直接與RNA結合,而是結合在其它蛋白質組分上。核糖體中的蛋白質,rRNA以及其他一些輔助因子在一起提供了翻譯過程所需的全部酶活性,這些酶活性只有在核糖體整體結構存在的情況下才具備。
不同來源的核糖體其形狀,大小、化學組成稍有不同,通常根據沉降係數的不同分為70S和80S兩種類型,70S核糖體存在於細菌,線粒體和葉綠體中,80s核糖體存在於真核生物的細胞質中。
所有的核糖體都是由大小兩個亞基構成,核糖體的大小亞單位只有在以mRNA為模板合成蛋白質時才結合在一起,肽鏈合成終止后,大小亞單位又解離,遊離於細胞質基質中。核糖體並不是單獨工作的,而是由多個甚至幾十個串連在一條mRNA分子上,稱多聚核糖體(polyribosome),這樣越長的mRNA可以結合更多的核糖體,提高了蛋白質合成的速度。
儘管原核生物與真核生物核糖體的蛋白質和rRNA差異很大,但結構總體相似,特別是負責與mRNA結合的小亞基。原核和真核細胞的rRNA都具有甲基化現象,這種甲基化與RNA的轉錄后加工過程的酶識別有關,另外原核5SrRNA和真核5.8SrRNA結構高度保守,常用於研究生物進化。