共找到5條詞條名為光合作用的結果 展開
光合作用
反應
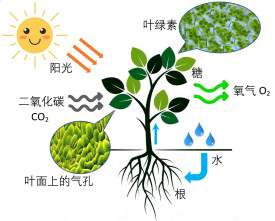
光合作用,通常是指綠色植物(包括藻類)吸收光能,把二氧化碳和水合成富能有機物,同時釋放氧氣的過程。其主要包括光反應、暗反應兩個階段,涉及光吸收、電子傳遞、光合磷酸化、碳同化等重要反應步驟,對實現自然界的能量轉換、維持大氣的碳-氧平衡具有重要意義。
綠色植物利用太陽的光能,同化二氧化碳(CO₂)和水(O₂)製造有機物質並釋放氧氣的過程,稱為光合作用。光合作用所產生的有機物主要是碳水化合物,並釋放出能量。
17世紀荷蘭科學家VanHelmont進行柳樹盆栽試驗。證明柳樹生長所需的主要物質不是來自土壤,而是來自水。
1771年英國牧師、化學家J.Priestley進行密閉鐘罩試驗,有植物存在蠟燭不熄滅,老鼠不會窒息死亡。1776年提出植物可以“凈化”空氣。1771年被稱為光合作用發現年。
1782年瑞士人JeanSnebier用化學方法發現CO₂是光合作用必需物質,O₂是光合作用產物。
1804年瑞士人N.T.DeSaussure做定量實驗證實植物所產生的有機物和所放出O₂的總量比消耗的CO₂多,證明還有水參與反應。
1864年J.V.Sachs發現照光葉片遇碘會變藍,證明光合作用形成碳水化合物(澱粉)。
19世紀末,證明光合作用原料是空氣中的CO₂和土壤中的H₂O,能源是太陽輻射能,產物是糖和O₂。
20世紀初,光合作用的分子機理有了突破性進展,里程碑式的工作主要是:Wilstatter等(1915)由於提純葉綠素並闡明其化學結構獲得諾貝爾獎。
隨後,英國的Blackman和德國的O.Warburg等人用藻類進行閃光試驗證明:光合作用可以分為需光的光反應(lightreaction)和不需光的暗反應(darkreaction)兩個階段。
1932年,R.Emersen和W.Arnold通過小球藻(chlorella)懸浮液做閃光試驗定義了“光合單位”(photosyntheticunit),即釋放1分子或同化1分子CO₂所需的2500個葉綠素的分子數目。隨後在1986年,Hall等人指出:光合單位應是包括兩個反應中心的約600個葉綠素分子(300×2)以及連結這兩個反應中心的光合電子傳遞鏈。而多數人贊同霍爾的看法,即:所謂的“光合單位”,就是指存在於類囊體膜上能進行完整光反應的最小結構單位。
1940年代~1950年代末,M.Calvin等用C研究光合碳同化,闡明了CO₂轉化為有機物的生化途徑。M.Calvin於1961年獲得諾貝爾獎。之後相繼確定了CAM途徑(M.Thomas,1960)和C4途徑(M.D.Hatch和C.B.Slack,1966)。
1954年,美國科學家D.I.Arnon等在給葉綠體照光時發現,當向體系中供給無機磷、和時,體系中就會有和產生。同時,只要供給和,即使在黑暗中,葉綠體也可將CO₂轉變為糖。因此,得出結論:光反應的實質在於產生“同化力”(assimilatorypower)去推動暗反應的進行,而暗反應的實質在於利用“同化力”將無機碳(CO₂)轉化為有機碳(CH₂O)。
1957年,Emerson觀察到小球藻在用遠紅光照射時補加了一點稍短波長的光(如650nm的光),則量子產額比這兩種波長的光單獨照射的總和還要高。這種在長波紅光之外再加上較短波長的光促進光合效率的現象被稱為雙光增益效應,或叫愛默生增益效應(Emersonenhancementeffect)。後來才知道,這是因為光合作用需要兩個光化學反應的協同作用。
1960年,Hill等人提出了雙光系統(twophotosystem)的概念,把吸收長波光的系統稱為光系統(photosystemⅠ,PSⅠ),吸收短波長光的系統稱為光系統Ⅱ(photosystemⅡ,PSⅡ),推動了PSⅠ和PS的分離、純化等生物化學與分子生物學的研究。
1965年,R.B.Woodward因全合成葉綠素分子等工作獲得了諾貝爾獎。
1980年代初,P.Mitchell提出化學滲透假說。Jagendorf等用葉綠體進行光合磷酸化分階段研究,證明光合磷酸化的高能態就是化學滲透假說中的跨膜質子梯度。這不僅使人們了解光合作用中能量轉換機制,並且導致將質子動力勢與離子運轉、類囊體結構動態變化和能量轉換反應調控過程聯繫起來研究。
1980年代末期,Deisenhofer等測定了光合細菌反應中心結構,取得了解膜蛋白複合體細節及光合原初反應研究的突出進展,獲得了1988年的諾貝爾獎。
1992年,Marcus因研究包括光合作用電子傳遞在內的生命體系的電子傳遞理論而獲得諾貝爾獎。
1990年代末,催化光合作用的光合磷酸化和呼吸作用的氧化磷酸化的酶的動態結構與反應機理研究獲得了重大進展。Walker和Boyer獲得了1997年的諾貝爾獎。
中國的光合作用研究自20世紀50年代開始,取得了長足的進展。如中國科學院上海植物生理研究所在光合作用能量轉換、光合碳代謝的酶學研究等方面,中國科學院植物研究所在光合作用的原初反應和光合色素蛋白複合體研究等方面都有所發現和創新。
儘管光合作用研究歷史不算長,但經過眾多科研工作者的努力探索,已取得了舉世矚目的進展,為指導農業生產提供了充分的理論依據。
光反應:
暗反應:
總反應:
其中表示糖類。
光反應階段的特徵是在光碟機動下水分子氧化釋放的電子通過類似於線粒體呼吸電子傳遞鏈那樣的電子傳遞系統傳遞給,使它還原為。電子傳遞的另一結果是基質中質子被泵送到類囊體腔中,形成的跨膜質子梯度驅動磷酸化生成。
暗反應階段是利用光反應生成NADPH和ATP進行碳的同化作用,使氣體二氧化碳還原為糖。由於這階段基本上不直接依賴於光,而只是依賴於ATP和NADPH的提供,故稱為暗反應階段。
| 反應階段 | 光反應 | 碳反應(暗反應) |
| 反應實質 | 光能→化學能,釋放 | 同化CO₂形成(CH₂O)(酶促反應) |
| 反應時間 | 短促,以微秒計 | 較緩慢 |
| 反應條件 | 需色素、光、ADP、和酶 | 不需色素和光,需多種酶 |
| 反應場所 | 在葉綠體內囊狀結構薄膜上進行 | 在葉綠體基質中進行 |
物質轉化 (光反應) | 2H₂O→4[H]+O₂↑ (在光和葉綠體中的色素的催化下) | CO₂+C₅→2C₃ (在酶的催化下) |
物質轉化 (暗反應) | ADP+Pi→ATP (在酶的催化下) | C₃+[H]→(CH₂O)+C₅ (在ATP供能和酶的催化下) |
| 能量轉化 | 葉綠素把光能先轉化為電能再轉化為活躍的化學能並儲存在ATP中 | ATP中活躍的化學能轉化變為糖類等有機物中穩定的化學能 |
植物在同化無機碳化物的同時,把太陽能轉變為化學能,儲存在所形成的有機化合物中。每年光合作用所同化的太陽能約為人類所需能量的10倍。有機物中所存儲的化學能,除了供植物本身和全部異養生物之用外,更重要的是可供人類營養和活動的能量來源。因此可以說,光合作用提供今天的主要能源。綠色植物是一個巨型的能量轉換站。
植物通過光合作用製造有機物的規模是非常巨大的。據估計,植物每年可吸收CO₂約合成約的有機物。地球上的自養植物同化的碳素,40%是由浮游植物同化的,餘下60%是由陸生植物同化的。人類所需的糧食、油料、纖維、木材、糖、水果等,無不來自光合作用,沒有光合作用,人類就沒有食物和各種生活用品。換句話說,沒有光合作用就沒有人類的生存和發展。
大氣之所以能經常保持21%的氧含量,主要依賴於光合作用(光合作用過程中放氧量約)。光合作用一方面為有氧呼吸提供了條件,另一方面,的積累,逐漸形成了大氣表層的臭氧(O3)層。臭氧層能吸收太陽光中對生物體有害的強烈的紫外輻射。植物的光合作用雖然能清除大氣中大量的CO₂,但大氣中CO₂的濃度仍然在增加,這主要是由於城市化及工業化所致。
光合速率通常是指單位時間單位葉面積所吸收的二氧化碳或釋放的氧氣的量,也可用單位時間單位葉面積上的干物質積累量來表示。
1.不同部位
在一定範圍內,葉綠素含量越多,光合越強。以一片葉子為例,最幼嫩的葉片光合速率低,隨著葉子成長,光合速率不斷加強,達到高峰,隨後葉子衰老,光合速率就下降。
2.不同生育期
株作物不同生育期的光合速率不盡相同,一般都以營養生長期為最強,到生長末期就下降。以水稻為例,分櫱盛期的光合速率較快,在稻穗接近成熟時下降。但從群體來看,群體的光合量不僅決定於單位葉面積的光合速率,而且很大程度上受總葉面積及群體結構的影響。
1.光照
(1)光強度對光合作用的影響
光合作用是一個光生物化學反應,所以光合速率隨著光照強庋的增減而增減。在黑暗時,光合作用停止,而呼吸作用不斷釋放CO₂;隨著光照增強,光合速率逐漸增強,逐漸接近呼吸速率,最後光合速率與呼吸速率達到動態平衡相等。同一葉子在同一時間內,光合過程中吸收的CO₂與光呼吸和呼吸作用過程中放出的CO₂等量時的光照強度,就稱為光補償點(lightcompensationpoint)。植物在光補償點時,有機物的形成和消耗相等,不能積累干物質,而晚間還要消耗干物質,因此從全天來看,植物所需的最低光照強度,必須高於光補償點,才能使植物正常生長。
(2)光質對光合作用的影響
光質也影響植物的光合效率。在自然條件下,植物會或多或少受到不同波長的光線照射例如,陰天的光照不僅光強弱,而且藍光和綠光成分增多;樹木的葉片吸收紅光和藍光較多,故樹冠下的光線富含綠光,尤其是樹木繁茂的森林更是明顯。
2.二氧化碳
二氧化碳是光合作用的原料,對光合速率影響很大。其主要是通過氣孔進入葉片,加強通風或設法增施二氧化碳能顯著提高作物的光合速率,對C₃植物尤為明顯。此外,植物對CO₂的利用與光照強度有關,在弱光情況下,只能利用較低濃度的CO₂,光合速率慢,隨著光照強度的加強,植物就能吸收利用較高濃度的CO₂,光合速率加快。
3.溫度
光合過程中的碳反應是由酶所催化的化學反應,而溫度直接影響酶的活性,因此,溫度對光合作用的影響也很大。除了少數的例子以外,一般植物可在10~35℃下正常地進行光合作用,其中以25~30℃最適宜,在35℃以上時光合作用就開始下降,40~50℃時即完全停止。在低溫中,酶促反應下降,故限制了光合作用的進行。而在高溫時,一方面是高溫破壞葉綠體和細胞質的結構,並使葉綠體的酶鈍化;另一方面,暗呼吸和光呼吸加強,光合速率便降低。
4.礦質元素
礦質元素直接或間接影響光合作用。氮鎂、鐵、錳等是葉綠素等生物合成所必需的礦質元素;銅、鐵、硫和氯等參與光合電子傳遞和水裂解過程;鉀、磷等參與糖類代謝,缺乏時便影響糖類的轉變和運輸,這樣也就間接影響了光合作用同時,磷也參與光合作用中間產物的轉變和能量傳遞,所以對光合作用影響很大。
5.水分
水分是光合作用原料之一,而光合作用所需的水分只是植物所吸收水分的一小部分(1%以下),因此,水分缺乏主要是間接地影響光合速率下降。具體來說,缺水使葉片氣孔關閉,影響CO₂進入葉內;缺水使葉片澱粉水解加強,糖類堆積,光合產物輸出緩慢,這些都會使光合速率下降。
● ● 葉片發育和結構1)葉齡:新長出的嫩葉,光合速率很低。不同部位葉片在不同生育期相對光合速率不同,光合速率隨葉齡增長出現“低-高-低”規律。2)葉結構:葉的結構,如葉厚度、柵欄組織與海綿組織的比例、葉綠體和類囊體的數目等都對光合速率有影響。葉的結構一方面受遺傳基因控制,另一方面則受環境影響。
● ● 光合產物的輸出光合產物(如蔗糖)從葉片輸出的速率會影響葉片的光合速率。形成原因:1)反饋抑制,2)澱粉粒的影響。
● ● 光照1)光強度對光合作用的影響光強度-光合速率曲線黑暗條件下,葉片不進行光合作用,只有呼吸作用釋放。隨著光強度的增加,光合速率也會相應提高;當到達某一特定光強度時,葉片的光合速率等於呼吸速率,即二氧化碳吸收量等於二氧化碳釋放量。當超過一定的光強,光合速率的增加就會轉慢。當達到某一光強時,光合速率不再增加,即光飽和點。光合作用的光抑制光照不足會成為光合作用的限制因素,光能過剩也會對光合作用產生不利影響。當光合機構接受的光能否超過所能利用的量時,會引起光合速率降低的現象。2)光質對光合作用的影響太陽輻射中,只有可見光部分才能被光合作用利用,光合作用的作用光譜與葉綠體色素的吸收光譜大體吻合。
● ● 二氧化碳1)二氧化碳-光合速率曲線二氧化碳是光合作用的原料,對光合速率影響很大。二氧化碳-光合速率曲線與光強曲線相似。2)二氧化碳的供給二氧化碳主要是通過氣孔進入葉片,加強通風或設法增施二氧化碳能顯著提高作物的光合速率,對碳三植物尤為明顯。
● ● 溫度光合過程的暗反應是由酶催化的生物化學反應,受溫度的強烈影響。
● ● 水分水分虧缺降低光合的主要原因有1)氣孔導度下降2)光合產物輸出變慢3)光合機構受損4)光合面積擴展受損
● ● 礦質營養礦物營養在光合作用中功能廣泛1)葉綠體結構的組成成分2)電子傳遞體的重要成分3)磷酸基團的重要作用4)活化或調節因子
● ● 光合速率的日變化一天中的環境因子在不斷變化,這些變化會使光合速率發生日變化,其中光強日變化對光合速率日變化影響最大。
植物在同化無機碳化物的同時,把太陽能轉變為化學能,儲存在所形成的有機化合物中。每年光合作用所同化的太陽能約為人能所需能量的10倍。
植物每年可吸收約合成約的有機物。。
主要依賴於光合作用(光合作用過程中放氧量約t/a)。光合作用一方面為有氧呼吸提供了條件,另一方面,的積累,逐漸形成了大氣表層的臭氧()層。臭氧層能吸收太陽光中對生物體有害的強烈的紫外輻射。植物的光合作用雖然能清除大氣中大量的,但大氣中的濃度仍然在增加,這主要是由於城市化及工業化所致。
基本信息
- 中文名
- 光合作用
- 外文名
- Photosynthesis
- 發生場所
- 綠色植物(包括藻類)的葉綠體、光合細菌
- 反應產物
- 富能有機物
- 環境影響
- 調節大氣碳-氧平衡
- 反應條件
- 光色素分子酶、二氧化碳(或硫化氫)