染色體
細胞核的組成部分
染色體(chromosome) 是細胞在有絲分裂或減數分裂時DNA存在的特定形式。細胞核內,DNA緊密卷繞在稱為組蛋白的蛋白質周圍並被包裝成一個線狀結構。
當細胞不分裂時,染色體在細胞核中是不可見的 - 在顯微鏡下也是如此。然而,構成染色體的DNA在細胞分裂過程中變得更緊密,在顯微鏡下可見。
每條染色體都有一個叫做著絲粒(點)的收縮點,它將染色體分成兩個部分,即“臂”。短臂為“p臂”;長臂為“q臂”。著絲粒(點)在每條染色體上的位置為染色體提供了特有的形狀,可用於幫助描述特定基因的位置。
染色體有種屬特異性,隨生物種類、細胞類型及發育階段不同,其數量、大小和形態存在差異。
染色體(chromosome)是真核細胞在有絲分裂或減數分裂時DNA存在的特定形式,由染色質絲螺旋纏繞,逐漸縮短變粗形成。
只有在細胞分裂中期(所有染色體以其濃縮形式在細胞中心排列),染色體通常在光學顯微鏡下才可見。在此之前,每個染色體已被複制一次(S階段),原來的染色體和其拷貝互稱姐妹染色體,兩個染色體通過著絲點(粒)連接。如果著絲點位於染色體的中間,則產生X形的染色體結構;如果著絲點位於其中一個末端附近,則產生雙臂的染色體結構。X形結構染色體被稱為中期染色體。在這種高度濃縮的形式中,染色體最容易區分和研究,易被鹼性染料(例如龍膽紫和醋酸洋紅)著色,因此而得名。
減數分裂過程中的染色體重組和隨後的有性繁殖在遺傳多樣性中發揮著重要作用。如果染色體不穩定性和易位發生的話,細胞有絲分裂將出現災難,細胞啟動細胞凋亡導致其自身死亡。細胞突變會阻礙這一過程,從而導致癌症發展。
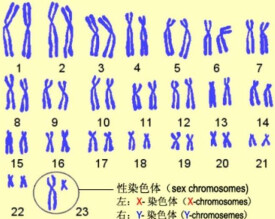
性細胞如精子、卵子等是單倍體,染色體數目只是體細胞的一半。哺乳動物雄性個體細胞的性染色體對為XY,雌性則為XX。鳥類。兩棲類。爬行類和某些昆蟲的性染色體與哺乳動物不同:雄性個體的是ZZ,雌性個體為ZW。
染色體(chromosome)來自希臘語χρῶμα(色度,“顏色”)和σῶμα(體細胞,“體”),描述了它們對特定染料的強染色。染色體由德國科學家von Waldeyer-Hartz創造,取代了發現細胞分裂的德國生物學家Walther Flemming提出的染色質(chromatin)。
1879年德國生物學家弗萊明(Fleming·w)把細胞核中的絲狀和粒狀的物質,用染料染紅,觀察發現這些物質平時散漫地分佈在細胞核中,當細胞分裂時,散漫的染色物體便濃縮,形成一定數目和一定形狀的條狀物,到分裂完成時,條狀物又疏鬆為散漫狀。
1883年美國遺傳學家、生物學家沃爾特·薩頓提出了遺傳基因在染色體上的學說。
1888年正式被命名為染色體。
1902年美國生物學家沃爾特·薩頓和鮑維里通過觀察發現細胞的減數分裂時染色體與基因具有明顯的平行關係,並推測基因位於染色體上。
1928年摩爾根通過果蠅雜交實驗證實了染色體是基因的載體,從而獲得了生理醫學諾貝爾獎。
1956年,美籍華裔遺傳學家Joe Hin Tjio(1919–2001,資料譯為庄有興或蔣有興)和Levan首次發現人的體細胞的染色體數目為46條,標誌著人類細胞遺傳學的建立。46條染色體按其大小、形態配成23對,第一對到第二十二對叫做常染色體,為男女共有,第二十三對是一對性染色體(Sex Chromosome),雄性個體細胞的性染色體對為XY;雌性則為XX。
序列結構
DNA包裝
原核生物不具有核,它們的DNA被組織在一個類核結構中。類核是獨特的結構並佔據細菌細胞確定的區域。但這種結構是動態的,可被與細菌染色體相關的一系列組蛋白樣蛋白的作用來維持和重塑。古細菌染色體中的DNA被包裝在與真核核小體相似的結構中。某些細菌還含有質粒或其它染色體外DNA。這些細胞質環狀結構含有細胞DNA,在水平基因轉移中發揮作用。細菌染色體傾向於結合於質膜上,因此,分子生物學通過離心裂解的細菌以便其從質粒DNA中的分離。與真核生物一樣,原核染色體和質粒DNA通常是超螺旋的。DNA轉錄、調節和複製前必須首先將DNA超螺旋鏈解開。
真核生物
真核生物中的染色體由染色質絲組成。染色質絲由核小體組成(組蛋白八聚體,DNA鏈的一部分附著並包裹在其周圍)。染色質絲被蛋白質包裝成稱為染色質的濃縮結構。染色質含有絕大多數的DNA和少量的母系遺傳獲得的如線粒體DNA。染色質存在於大多數細胞中,除少數例外,例如紅細胞。染色質允許非常長的DNA分子進入細胞核。在細胞分裂期間,染色質進一步濃縮以形成顯微鏡下可見的染色體。染色體的結構隨細胞周期而變化。在細胞分裂期間,染色體被複制,分裂並成功傳遞給它們的子細胞,以確保它們的後代的遺傳多樣性和存活。染色體可以以複製或非複製的形式存在。未複製的染色體是單個雙螺旋,而複製的染色體包含由著絲粒連接的兩個相同的雙螺旋(其中的任何一個被稱為染色單體或姐妹染色單體)。
真核生物(具有細胞核的細胞,例如植物、真菌和動物細胞)具有包含在細胞核中的多個大的線性染色體。每個染色體都有一個著絲粒,一個或兩個從著絲點突出的臂。此外,大多數真核生物還有小的環狀線粒體染色體,一些真核生物也有額外的小環狀或線性細胞質染色體。在真核生物的核染色體中,未濃縮的DNA以半有序結構存在,被包裹在組蛋白(結構蛋白)周圍,形成稱為染色質的複合結構。
間期染色質
在細胞不分裂的間期,存在兩種類型的染色質:常染色質,由具有活性的DNA組成;異染色質,主要由無活性的DNA組成,似乎在染色體階段起到結構性作用。異染色質可進一步區分為兩種類型:組成型異染色質,位於著絲粒周圍,通常包含重複序列,從未表達;兼性異染色質,有時表達。
中期染色質
在有絲分裂或減數分裂(細胞分裂)的早期,染色質雙螺旋變得越來越濃縮。此時的染色體不再是可以進入的遺傳物質(轉錄停止),而是一種緊湊的可運輸的結構,形成經典的四臂結構,一對姐妹染色單體在著絲粒處相互連接。較短的手臂被稱為p臂,較長的手臂稱為q手臂。這個時期是用光學顯微鏡觀察單個染色體的最佳時間。有絲分裂中期的染色體是線性的縱向壓縮的連續的染色質組成的環。在有絲分裂期間,微管從著絲點生長,並且通過被稱為動粒的特殊結構附著到著絲點上。每個染色單體具有自己的動粒,反向附著於有絲分裂紡錘體的兩極。在從中期到後期的過渡之中,微管將染色單體拉向著絲粒,使每個子細胞繼承一組染色單體。一旦細胞分裂,染色單體就會被解開,DNA可以再次被轉錄。染色體結構上的高度濃縮,使得這些巨大的DNA結構能夠包含在細胞核內。
人類染色體可分為兩種類型:常染色體(體染色體)和性染色體(異體染色體)。某些遺傳特徵與一個人的性別有關,並通過性染色體傳播。常染色體因此包含其餘部分的遺傳信息。常染色體和性染色體的複製、有絲分裂和減數分裂過程一致。
人類細胞有23對染色體(22對常染色體和一對性染色體),即每個細胞共有46個染色單體。除此之外,人類細胞還有數百個線粒體染色體拷貝。人類基因組的測序提供了關於每條染色體的大量信息。下表是根據Sanger Institute在脊椎動物基因組註釋(VEGA)資料庫中的人類基因組信息編製的染色體統計數據。基因數量是估計值,因為它部分基於基因預測。總染色體長度也是估計值,是基於未測序異染色質區域的大小估計的。
| 染色體 | 基因 | 總的鹼基對 | % (鹼基) | 測序的鹼基對 | % (測序的鹼基對) |
|---|---|---|---|---|---|
| X (sex chromosome) | 800 | 154,913,754 | 5.0 | 151,058,754 | 97.51% |
| Y (sex chromosome) | 200 | 57,741,652 | 1.9 | 25,121,652 | 43.51% |
| 1 | 2000 | 247,199,719 | 8.0 | 224,999,719 | 91.02% |
| 2 | 1300 | 242,751,149 | 7.9 | 237,712,649 | 97.92% |
| 3 | 1000 | 199,446,827 | 6.5 | 194,704,827 | 97.62% |
| 4 | 1000 | 191,263,063 | 6.2 | 187,297,063 | 97.93% |
| 5 | 900 | 180,837,866 | 5.9 | 177,702,766 | 98.27% |
| 6 | 1000 | 170,896,993 | 5.5 | 167,273,993 | 97.88% |
| 7 | 900 | 158,821,424 | 5.2 | 154,952,424 | 97.56% |
| 8 | 700 | 146,274,826 | 4.7 | 142,612,826 | 97.50% |
| 9 | 800 | 140,442,298 | 4.6 | 120,312,298 | 85.67% |
| 10 | 700 | 135,374,737 | 4.4 | 131,624,737 | 97.23% |
| 11 | 1300 | 134,452,384 | 4.4 | 131,130,853 | 97.53% |
| 12 | 1100 | 132,289,534 | 4.3 | 130,303,534 | 98.50% |
| 13 | 300 | 114,127,980 | 3.7 | 95,559,980 | 83.73% |
| 14 | 800 | 106,360,585 | 3.5 | 88,290,585 | 83.01% |
| 15 | 600 | 100,338,915 | 3.3 | 81,341,915 | 81.07% |
| 16 | 800 | 88,822,254 | 2.9 | 78,884,754 | 88.81% |
| 17 | 1200 | 78,654,742 | 2.6 | 77,800,220 | 98.91% |
| 18 | 200 | 76,117,153 | 2.5 | 74,656,155 | 98.08% |
| 19 | 1500 | 63,806,651 | 2.1 | 55,785,651 | 87.43% |
| 20 | 500 | 62,435,965 | 2.0 | 59,505,254 | 95.31% |
| 21 | 200 | 46,944,323 | 1.5 | 34,171,998 | 72.79% |
| 22 | 500 | 49,528,953 | 1.6 | 34,893,953 | 70.45% |
| Total | 21,000 | 3,079,843,747 | 100.0 | 2,857,698,560 | 92.79% |
真核生物
下面這些表給出了真核生物細胞核中染色體(包括性染色體)的總數。大多數真核生物是二倍體,如人具有22種常染色體(每個有兩個同源拷貝)和1種性染色體(2個拷貝),總共有46條染色體。其它生物染色體有兩個以上的拷貝,例如麵包小麥,它是六倍體,有七種不同染色體,各為六個拷貝,總共42個染色體。
| 常見生物染色體數目列表 | ||
| 通名 | 學名 | 雙倍體數 |
|---|---|---|
| 動物 | ||
| 人類 | Homo sapiens | 46 |
| 彌猴 | Macacamalatta | 42 |
| 黃牛 | Bostaurus | 60 |
| 豬 | Susscrofa | 38 |
| 狗 | Canis familiaris | 78 |
| 貓 | Felis domesticus | 38 |
| 馬 | Equus Calibus | 64 |
| 驢 | Equus asinus | 62 |
| 山羊 | Capara hircus | 60 |
| 綿羊 | Ovis aries | 54 |
| 小家鼠 | Mus musculus | 40 |
| 大家鼠 | Rattus norvegicus | 42 |
| 水貂 | Mustela vison | 30 |
| 豚鼠 | Cavia cobaya | 64 |
| 兔 | Oryctolagus cuniculus | 44 |
| 家鴿 | pigeon | 約80 |
| 雞 | Gallus domesticus | 約78 |
| 火雞 | Meleagris gallopavo | 約80 |
| 鴨 | Anas platyrhyncho | 約80 |
| 家蠶 | Bombyx mori | 56 |
| 家蠅 | Musca domestica | 12 |
| 果蠅 | Drosophila melanogaster | 8 |
| 蜜蜂 | Apis mellifera | ♀32♂16 |
| 蚊 | Culex pipiens | 6 |
| 佛蝗 | Phlaeobainfumata | ♀24♂23 |
| 水螅 | hydra | 32 |
| 植物 | ||
| 洋蔥 | Alliumcepa | 16 |
| 大麥 | Hordeum uulgare | 14 |
| 水稻 | Oryza sativa | 24 |
| 小麥 | Triticum vulgare | 42 |
| 玉米 | Zea mays | 20 |
| 金魚草 | Antirrhinum majus | 16 |
| 陸地棉 | Gossypium hirsutum | 52 |
| 中棉 | Gossypium arboreum | 26 |
| 豌豆 | Pisum sativum | 14 |
| 香豌豆 | Lathyrus odoratus | 14 |
| 蠶豆 | Vicia faba | 12 |
| 菜豆 | phaseolus vulgaris | 22 |
| 向日葵 | Helianthusannuus | 34 |
| 煙草 | Nicotiana taldcum | 48 |
| 番茄 | Solanum lycopersicum | 24 |
| 松 | Pinus species | 24 |
| 青菜 | Brassica chinensis | 20 |
| 甘藍 | Brassica oleracea | 18 |
| 月見草 | Oenothera biennis | 14 |
| 微生物 | ||
| 鏈孢霉 | Neurospora crassa | 7 |
| 青黴菌 | Penicillium species | 4 |
| 麴黴 | Aspergillus nidulans | 8 |
| 衣藻 | Chlamydomonas reinhardi | 16 |
每個真核生物種類的不同成員都具有相同數量的核染色體(見表),真核生物的其它染色體,如線粒體和質體染色體數量的差異較大,每個細胞可能有數千個拷貝。
無性繁殖物種所有體細胞中都具有一套相同的染色體,但無性繁殖物種可以是單倍體或二倍體。有性繁殖物種的體細胞是二倍體,有兩套染色體,一套來自母親,一套來自父親。配子即生殖細胞是單倍體:它們只有一套染色體。配子由二倍體生殖細胞減數分裂產生。減數分裂過程中,父母匹配的染色體可以通過交會發生一小部分遺傳物質的交換,從而產生的新染色體不僅僅只含有從父母方繼承來的遺傳信息。當雄雌配子結合(受精)後會形成新的二倍體生物。
一些動植物物種是多倍體。它們具有兩組以上的同源染色體。與其祖先物種相比,重要的農業植物如煙草或小麥通常是多倍體。野生小麥二倍體含14個染色體,硬質小麥和麵包小麥是多倍體,分別有28個(四倍體)和42個(六倍體)染色體。
原核生物
原核生物通常只含有一個主要染色體,但大多數多拷貝的細胞也很容易存活。例如大型細菌Epulopiscium fishelsoni中存在多達10萬個拷貝的染色體。與真核生物一樣,質粒和質粒樣小染色體的拷貝數數目差異較大。細胞中質粒數量幾乎完全由質粒分裂速率決定,分類快,拷貝數多。
核型變異
一般而言,核型是真核生物物種特徵性染色體的補體。核型的製備和研究是細胞遺傳學的一部分。
儘管真核生物DNA複製和轉錄是高度標準化,但它們的核型通常是高度可變的。物種之間的染色體數目和組織定位可能存在差異。在某些情況下,物種內也存在顯著差異。通常有:
1)兩性之間的差異
2)體細胞和生殖細胞之間的變異
3)遺傳多態性造成的群體成員之間的差異
4)種族之間存在的地域差異
5)複合原因或其它個體異常
此外,受精卵的發育過程異常也可能導致核型的變化。
核型分析
確定核型的技術通常稱為核型分析。細胞可以秋水仙鹼體外固定分裂(中期)細胞,然後將這些細胞染色,拍照並按照先常染色體(長度順序)后性染色體(此處為X / Y)的順序將染色體排列成核型。像許多有性生殖的物種一樣,人類有特殊的性染色體,女性為XX,男性為XY。
染色體易位或染色體倒位不會在攜帶者中引起疾病,但它們可能提高其後代發病的機會。
染色體或染色體組的數目異常,又稱為非整倍體,可能是致命的,或可能導致遺傳性疾病
基本信息
- 中文名
- 染色體
- 外文名
- Chromosome
- 細分類型
- 常染色體、性染色體(包含X染色體和Y染色體)
- 意義
- 遺傳信息的載體
- 名稱來源
- 易被鹼性染料 如龍膽紫 醋酸洋紅著色
- 組成
- 主要為DNA和蛋白質