基因重排
基因重排
基因重排是指將一個基因從遠離啟動子的地方移到距啟動子很近的地方從而啟動轉錄的方式。
基因重排分基因內重排和基因間重排。
基因結構重排的機制是一種DNA雙鏈斷裂(double-stand break)的修復過程,在等位基因內或等位基因之間,出現了重複單位複雜的轉換式移動( conversional transfer)。
DNA雙鏈斷裂常發生在靠近串聯重複序列的5’端的重複單位內,形成DNA分子的兩個遊離的、突出的單鏈末端。但是在修復的過程中,兩末端因擺動或錯位,沒有按照原配對鹼基的位置復性,出現了兩種後果:
一個結果是錯位鏈最末端的鹼基率先復性,然後局部合成空缺的鹼基,經過修復形成一個或幾個插入重複單位。因為是發生在同- DNA分子內的單鏈插入,故這種基因的轉移是一種基因內轉換形式。基因內轉換重排可以反覆出現,每出現一次就增加一段插入序列,所以這種錯位復性及修復方式在小衛星座位一般都是增加了重複單位數,如圖表示重複單位由原來的7個重複單位突變增加到9個重複單位。
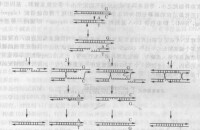
基因間重排
(1)從重複序列開始的錯配新合成鏈,多數在達到重複序列的側翼序列之前就被DNA錯配修復系統(mismatch repair system)終止,只能形成基因插入轉換。圖所示侵入鏈被錯配修復體系終止,又回到原單鏈繼續複製過程。此時,該單鏈上已經插入了來自對應的另一姐妹染色單體的3個重複單位,由原來7個重複單位突變為10個重複單位。這種被修復系統監測並終止的DNA雜合雙鏈可能再次分離,開始新一輪的鏈侵入、合成和單鏈復性,引起雜合鏈的再次延長。最後單鏈缺口被填充,雜合雙鏈區被修復。這種修復后的重排方式最多見。
(2)第二種是以對應同源染色單體的等位基因為模板合成的錯配鏈一直延伸到小衛星重複序列的側翼區,並連同側翼序列中可能存在的SNP基因座的鹼基變異一起發生了基因間轉換,這種重排同時涉及小衛星重複單位和側翼序列SNP基因座,因此稱為基因共轉換(coconversion)。共轉換的方式相對較少。圖2示侵入鏈以對應同源染色單體為模板延伸至側翼區的SNP座位,獲得SNP座位的C鹼基。被修復系統終止后,再回到原來的單鏈按原來序列繼續複製,此時新鏈的側翼SNP座位已經是C/G而不是原來的A/T。
(3)第三種結果更少見,即延伸的雜合雙鏈沒有終止,越過串聯重複區段,最後形成典型的Holliday連接結構,然後出現基因間重組交換(crossover)的過程,經過不同的拆分方式形成同源染色單體之間的互換產物,如圖所示。
對CEBl (D2S90)小衛星的研究發現,在配子細胞中,基因轉換和重排引起的突變率要比傳統概念的配子不等交換突變高70倍,因此認為基因重排是小衛星的多態性形成的主要原因。男性配子基因轉換突變率遠高於女性配子,試驗數據顯示男性配子突變率13%,女性僅o.4%。對小衛星和側翼SNP標記的基因共轉換觀察,也發現轉換事件與側翼SNP的某種單倍型有明顯的關聯,推測小衛星序列的基因轉換重排可能受到側翼DNA序列的調節。
基因組重排技術結合了傳統誘變技術和細胞融合技術,是一項對整個微生物基因組重排的新型育種技術。基因組重排技術通過多親本原生質體遞歸融合,可以使工程菌快速獲得多樣複雜優良表型,並且無須了解其基因組學、代謝組學等具體背景。介紹了基因組重排技術的過程及應用,展現了基因組重排技術的優點,並給出了基因組重排技術的發展在未來的應用情景。
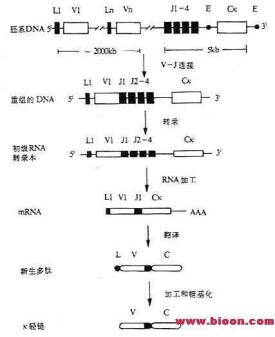
另外,在B淋巴細胞發育過程中,也有關於基因重排的機制。
基本信息
- 中文名
- 基因重排
- 分類
- 基因內重排和基因間重排
- 應用
- 新型育種技術
- 內容
- 移動基因到啟動子附近