樹突狀細胞
抗原提呈細胞之一
樹突狀細胞是由加拿大學者Steinman於1973年發現的,是目前所知的功能最強的抗原提呈細胞,因其成熟時伸出許多樹突樣或偽足樣突起而得名。
加拿大學者Steinman。
樹突狀細胞
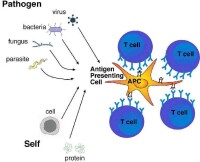
樹突狀細胞(dendritic cells, DC)廣泛分佈於腦以外的全身組織和臟器,數量較少,僅占人外周血單個核細胞的1%,因其具有許多分枝狀突起故名。根據來源,可將DC 分為兩類,即來源於髓系幹細胞的髓樣樹突狀細胞(myeloid DC)和來源於淋巴系幹細胞的淋巴樣樹突狀細胞(lymphoid DC)。這些DC 因其分佈情況或分化程度的不同而有不同的名稱。例如,位於表皮和胃腸上皮組織中的DC 稱為朗格漢斯細胞(langerhan's cells,LC);心、肺、肝、腎等器官結締組織中的DC 稱為間質樹突狀細胞(interstitial DC );外周免疫器官胸腺依賴區和胸腺髓質區的DC 稱為並指樹突狀細胞(interdigitating cell ,IDC);外周免疫器官淋巴濾泡區的DC 稱為濾泡樹突狀細胞( follicular dendritic cells,FDC);淋巴液中的DC 稱為隱蔽細胞(veiled cell)。在上述DC 中,除胸腺髓質區的DC 屬淋巴系DC 外,在末梢組織器官中的DC 屬髓系或淋巴系DC。其中位於表皮和胃腸上皮組織中的朗格漢斯細胞和位於實體器官結締組織中的間質DC 屬未成熟DC。這些未成熟DC 在接受抗原或炎性介質等作用刺激后,可發育分化為成熟的DC。樹突狀細胞是專職抗原提呈細胞,其主要功能是攝取、加工處理和提呈抗原,啟動特異性免疫應答。未成熟DC 可表達MHCⅠ/Ⅱ類分子、 IgGFc 受體(FcγR)、C3b 受體(C3bR)和某些Toll 樣受體如TLR2、TLR4 及TLR9。未成熟DC 攝取、加工處理抗原能力強,而提呈抗原激發免疫應答能力弱。成熟DC 表面特徵性標誌為CD1a、CD11c 和cd83,可高表達MHC-Ⅱ/Ⅰ類分子和共刺激分子(如B7 和ICAM),其攝取、加工處理抗原能力弱,而提呈抗原、啟動免疫應答能力強。DC 能誘導初始T 細胞活化,因此是機體特異性免疫應答的始動者。樹突狀細胞也是體內重要的免疫調節細胞,可通過分泌不同的細胞因子參與固有和適應性免疫應答。例如:①有些DC 可分泌以IL-12 為主的細胞因子,誘導或促進初始T 細胞分化為Th1 細胞,增強細胞免疫應答;②有些DC 可分泌以I 型干擾素為主的細胞因子,產生抗感染和免疫調節等作用;③在有些情況下,DC 可通過分泌IL-10 和TGF-β 等細胞因子,誘導B 細胞發生Ig 類別轉換,產生 IgA 類抗體;也可通過分泌以IL-1β為主的細胞因子,促進T、B 細胞活化。
人樹突狀細胞起源於造血幹細胞(hemopoieticstemcell)。DC的來源有兩條途徑:
①髓樣幹細胞在GM-CSF的刺激下分化為DC,稱為髓樣DC(myeloid dendritic cells,MDC),也稱DCl,與單核細胞和粒細胞有共同的前體細胞;包括朗格漢斯細胞(Langerhans cells ,LCs),間皮(或真皮)DCs以及單核細胞衍生的DCs等。
②來源於淋巴樣子細胞,稱為淋巴樣DC(Lymophiod dendritic cells,LDC)或漿細胞樣DC(plasmacytoid dendritic cells,piX;),即DC2,與T細胞和NK細胞有共同的前體細胞。樹突狀細胞(DC)儘管數量不足外周血單核細胞的1%,但表面具有豐富的抗原遞呈分子(MHC-Ⅰ和MHC-Ⅱ)、共刺激因子(CD80/B7-1、CD86/B7-2、CD40、CD40L等)和粘附因子(ICAM-1、ICAM-2、ICAM-3、LFA-1、LFA-3等),是功能強大的專職抗原遞呈細胞(APC)。DC自身具有免疫刺激能力,是惟一能激活未致敏的初始型T細胞的APC。
人體內大部分DC處於非成熟狀態,表達低水平的共刺激因子和粘附因子,體外激發同種混合淋巴細胞增殖反應的能力較低,但未成熟DC具有極強的抗原吞噬能力,在攝取抗原(包括體外加工)或受到某些因素刺激時即分化為成熟DC,而成熟的DC表達高水平的共刺激因子和粘附因子。DC在成熟的過程中,由接觸抗原的外周組織遷移進入次級淋巴器官,與T細胞接觸並激發免疫應答 樹突狀細胞(DC)儘管數量不足外周血單個核細胞的1%,但表面具有豐富的抗原遞呈分子(MHC-Ⅰ和MHC-Ⅱ)、共刺激因子(CD80/B7-1、CD86/B7-2、CD40、CD40L等)和粘附因子(ICAM-1、ICAM-2、ICAM-3、LFA-1、LFA-3等),是功能強大的專職抗原遞呈細胞(APC)。DC自身具有免疫刺激能力,是目前發現的惟一能激活未致敏的初始型T細胞的APC。
人體內大部分DC處於非成熟狀態,表達低水平的共刺激因子和粘附因子,體外激發同種混合淋巴細胞增殖反應的能力較低,但未成熟DC具有極強的抗原吞噬能力,在攝取抗原(包括體外加工)或受到某些因素刺激時即分化為成熟DC,而成熟的DC表達高水平的共刺激因子和粘附因子。DC在成熟的過程中,由接觸抗原的外周組織遷移進入次級淋巴器官,與T細胞接觸並激發免疫應答。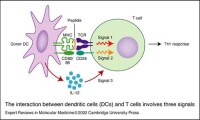
樹突狀細胞
DC作為目前發現的功能最強的APC,能夠誘導特異性的細胞毒性T淋巴細胞(cytotoxic T lymphocyte,CTL)生成。近年來研究表明,應用腫瘤相關抗原或抗原多肽體外衝擊致敏DC,回輸或免疫接種於載瘤宿主,可誘發特異性CTL的抗腫瘤免疫反應。
DC與腫瘤的發生、發展有著密切關係,大部分實體瘤內浸潤的DC數量多則患者預后好。有效的抗腫瘤免疫反應的核心是產生以CD8+ T細胞為主體的細胞免疫應答,這也是DC作為免疫治療手段的基礎。
DC抗腫瘤的機制如下:①DC可以高表達MHC-Ⅰ類和MHC-Ⅱ類分子,MHC分子與其捕獲加工的腫瘤抗原結合,形成肽-MHC分子複合物,並遞呈給T細胞,從而啟動MHC-I類限制性CTL反應和MHC-Ⅱ類限制性的CD4+ Thl反應。同時,DC還通過其高表達的共刺激分子(CD80/B7-1、CD86/B7-2、CD40等)提供T細胞活化所必須的第二信號,啟動了免疫應答。②DC與T細胞結合可大量分泌IL-12、IL-18激活T細胞增殖,誘導CTL生成,主導Th1型免疫應答,利於腫瘤清除;激活穿孔素P顆粒酶B和FasL/Fas介導的途徑增強NK細胞毒作用;③DC分泌趨化因子(Chemotactic Cytokines, CCK)專一趨化初始型T細胞促進T細胞聚集,增強了T細胞的激發。保持效應T細胞在腫瘤部位長期存在,可能通過釋放某些抗血管生成物質(如IL-12、IFN-γ)及前血管生成因子而影響腫瘤血管的形成。上述CCK進一步以正反饋旁分泌的方式活化DC,上調IL-12及CD80、CD86的表達;同時DC 也直接向CD8+T細胞呈遞抗原肽,在活化的CD4+ T細胞輔助下使CD8+ T細胞活化,CD4+ 和CD8+T細胞還可以進一步通過分泌細胞因子或直接殺傷,增強機體抗腫瘤免疫應答。
樹突狀細胞
在一段成長過程中他們長出樹枝狀的突起,原文dendrite來自希臘文的dentrites(關於樹的),因此本文譯為樹突狀的細胞。然而,這些並不與神經元有特殊的關聯,雖然其也有相似的部位。未成熟的樹突狀細胞也叫做隱匿性細胞,不若樹枝狀的突起,它們具有大量的、細胞質的菌幕。
基本信息
- 發現時間
- 1973年
- 作用
- 啟動、調控、並維持免疫應答中心
- 發現人
- 加拿大學者Steinman
- 中文名
- 樹突狀細胞
- 外文名
- Dendritic cells, DC