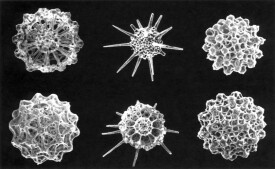
放射蟲
輻足綱下屬亞綱原生動物的通稱
原生動物門輻足綱中的等輻骨亞綱稀孔亞綱和多囊亞綱原生動物的通稱。因偽足和骨骼大都呈輻射狀而得名。放射蟲種類多,數量大,死亡后沉積海底所形成的軟泥占現代海底面積的2~3%(僅次於有孔蟲)。成蟲有單體或群體,大小相差懸殊,由幾微米至數毫米,某些群體的直徑可達數米,但絕大多數是單細胞的個體在此處添加文字。

放射蟲
分類位置:原生動物門輻足綱放射蟲亞綱
主要特徵:在細胞質內有一個球形、梨形或圓盤形的幾丁質中心囊。中心囊將細胞質分成囊外和囊內兩部分。囊的表面包以角質膜,膜上有小孔,使囊內外的細胞質相互溝通。囊內有核,司營養及生殖;囊外細胞質常有許多脂肪粒和空泡,有利於動物的漂浮生活。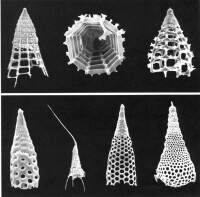
放射蟲
生態習性:大洋環境,營漂浮生活
分佈:廣泛,遍及所有海域。溫帶地區,尤其是赤道地區最為豐富。
具有結構精緻的硅質骨骼,發育著名的放射蟲軟泥。
地史分佈:寒武紀——現代
繁盛期:D、C、J、K、始新世、中新世,現代為極盛。
已知化石放射蟲2000種以上,現生放射蟲6000種以上。
第一階段:
十九世紀初,蘇俄的一名水手第一次發現放射蟲。
第二階段:
十九世紀中葉——末期
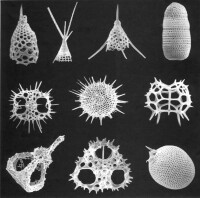
放射蟲
3、Ehrenberg(1830-1875):第一個廣泛研究放射蟲的學者。《Microgeologie》記述、定義了采自地中海的放射蟲,提議用Radiolaria來代表呈輻射對稱的海生肉足類。
4、Haeckel(1862):修訂Radiolaria包括具中心囊的輻射對稱的海生肉足類,並進行了最初的分類。描述了430种放射蟲,新種350多種(挑戰者號1872--1876)
5、十九世紀末期歐洲和俄國也發表了有關報道。
6、K. Zittel(德)最早從岩石中發現放射蟲化石。
7、Ehrenberg從第三紀硅質岩中發現了放射蟲化石
第三階段:
二十世紀七十年代——現代
深海鑽探計劃(JOIDES)、板塊構造理論促使放射蟲研究再度萌芽並蓬勃發展。主要在放射蟲的形態學、生態學和生物地層學方面進行了系統的研究,取得了重要的進展。美國加州海洋學院——Prof. W. R.ReiedelUniv. of Texas at Dallas——Prof.E.A.Pessagno,Jr. (放射蟲之父)
我國研究歷史
台灣地區八十年代後期起步
吳浩若、楊群到Dallas學習,研究地區為 海洋、造山帶
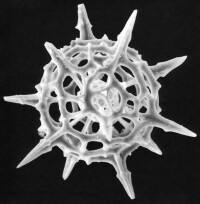
放射蟲
1、個體大小
單體:成年個體直徑數十µm~數百 µm
群體:直徑數厘米~數十厘米
2、生活方式:浮遊
單細胞為大多數
群體生活:泡沫蟲中膠球蟲科的一些種,單體由膠質相連,外包有半透明的膠質包膜
3、原生質構造
中心囊:球形、鐘罩形、葉狀或不規則形。囊壁是一層多孔的假幾丁質或類粘蛋白質的膜。放射蟲的骨架主要隨中心囊的形狀架構而成。
中心囊的構造差異是分類上極為重要的依據。
泡沫蟲:中心囊呈軸對稱,囊壁為單層的薄膜,具無數規則分佈的小孔。
罩籠蟲:中心囊呈狹長型,單軸式的放射狀對稱,囊壁為一層緻密的薄膜,僅在長軸的一端有孔洞。
囊內質:一個或數個細胞核,複核、各種細胞類器官、脂肪球、色素體、結晶體,司營養和生殖功能。
囊外質:以多囊蟲目為例
肉基質層:顆粒狀原生質組成,軸偽足由此伸出,消化作用在此進行,常可見半消化物質。
膠泡層:厚層的膠狀組織,充滿膠泡,可調節比重,利於漂浮,分泌骨骼。蟲黃藻共生於此。
肉網層:外質的最外側,網目狀構造,周邊向外延伸出絲狀偽足。
偽足:偽足上可附有粘稠狀的硅膠。軸狀偽足:由微管組成的軸絲支持;絲狀偽足
二、營養與繁殖
1、營養
放射蟲主要靠偽足捕食各種浮游生物。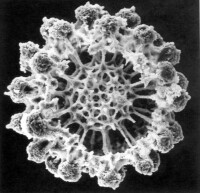
放射蟲
共生的藻類(蟲黃藻)也可為放射蟲提供營養。僅限於透光帶。共生的藻類散居在膠泡層中也有在肉基質層附近或聚集在中心囊內。共生的藻類也可成為放射蟲的食物。
2 、生活周期
很短,數天至一個月左右
3、繁殖
無性生殖(分裂生殖、出芽生殖),有性生殖。群體放射蟲有大量具鞭毛的細胞。雙型現象(Kling,1970)
三、骨骼
大多數的放射蟲具有結構精緻的硅質骨架,但海水表層常見的一種群居放射蟲就沒有骨架(Collozoum inerme,膠體放射蟲)。
1、骨骼成分與特性
主要為Si和O,還有少量的等元素,總量不超過4%。骨骼清晰透明,透射光下、呈玻璃狀,硬且脆,無彈性。
A:非晶硅質(蛋白石質)——多囊蟲目
放射蟲
骨骼成分相當穩定,放射蟲化石都是蛋白石質。
B:有機質和蛋白石混合成分——褐囊蟲目
沉積過程中被分解,無法形成化石,在海底表層沉積物中偶見。
C:硫酸鍶(天青石)——棘刺蟲,從未發現化石
2、骨骼類型
鬆散結構:為互不連接或接合不堅實的桿、骨針及刺
網格狀:孔的大小和形狀是重要的鑒定特徵
海綿狀: 細短的小棒三維空間連接。
孔板狀: 緻密均勻,可見稀疏、大小不等的孔。
3、骨骼形態
A:泡沫蟲類:球形,有多個同心球殼構成,球殼之間有放射狀小梁相連,放射狀刺從球體表面伸出。分為髓殼和皮殼
B:罩籠蟲類:一極開口的異極殼,軸對成或兩側對稱
C:阿爾拜蟲類:兩側對稱,殼壁多為無孔板狀
4、骨骼發育
Haeckel的兩種假說:其一骨骼單元直接由偽足和原生質網的化學成分變質形成;其二由原生質分泌形成。Thompson用在細胞質分界面上發生的物理-化學過程來解釋骨骼形成的網狀模式Hollande認為骨骼形成於活的原生質膜內。個體發育的不同階段,骨骼的形態和結構發生了 一系列的變化。在成年以後,骨骼還具有次生生長現象,表現為翼膜和附加骨骼網。
5、骨骼保存
與保存的環境有關,0-1000米貧硅水體;火山活動有利於放射蟲的生活和保存,有機組分沉積速率高的地方易保存。CCD面以下,常成為沉積物的主要成分。放射蟲軟泥,主要分佈於熱帶太平洋和印度洋深海底。與放射蟲骨骼本身的結構有關,骨骼纖細的種類更易溶解。
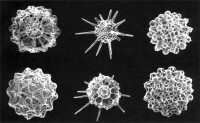
放射蟲
放射蟲的分佈
1、水平分佈
外海,遠洋,在淺海外陸架有少量分佈。
2、垂直分佈
可生活在不同的水深,海水表面至數千米深的洋底。表層個體較小,深海水域中個體較大。大多數生活在表層透光帶內(0—幾百米),表層、中層上部——棘刺蟲類、泡沫蟲類和一些罩籠蟲類;中層以下——一些罩籠蟲類、褐囊蟲類
溫度:
多數放射蟲分佈在溫暖的海洋中,由赤道向兩極數量很快減少。僅很小部分可在兩極生存。在大洋表層可劃分出四個組合——極區帶、近極帶、亞熱帶和熱帶表層組合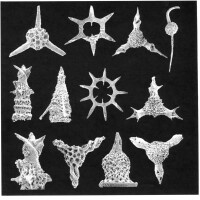
放射蟲
寒流地區——放射蟲種類少,殼壁厚,構造緻密,刺短而粗
暖流地區——放射蟲種類多,殼壁薄,構造纖細,孔小,刺長,側棘發育。
深度:
各組合的深度限制界限位於50,200,400,1000和4000米
鹽度:
放射蟲對鹽度的反應較為靈敏,高於溫度。部分類群可生活於極區,陸地淡水沒有發現。正常鹽度種類豐富。
水團:
是放射蟲生存的環境。同一水平面分佈不同的水團,垂向的不同層次具有不同的水團。水團的性質決定放射蟲組合特徵的差異。北極從表層到深處只有一個水團低緯度地區可具有不同的水團。
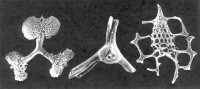
放射蟲
1、棘刺蟲亞目(Acantharia)
2、泡沫蟲亞目(Spumellina)
3、罩籠蟲亞目(Nassellina)
4、褐囊蟲亞目(Phaeodarina)
C.Deflandre建立阿爾拜蟲亞目(Albaillellaria)
Class Actinopoda Calkins 輻足蟲綱:具有軸偽足的肉足動物,廣義的放射蟲類。
Subclass Acantharia Haeckel 棘刺蟲亞綱
Subclass Radiolaria Müller 放射蟲亞綱
褐囊蟲目(Phaeodaria)
多囊蟲目(Polycystina)
泡沫蟲亞目(Spumellina)
罩籠蟲亞目(Nassellina)
阿爾拜蟲亞目(Albaillellaria)
Subclass Acantharia Haeckel 棘刺蟲亞綱
大多數種具中心囊膜,其上均勻分佈許多小孔。骨骼產生於細胞中心,類型多樣。骨骼成分主要為硫酸鍶。
中心囊內常共生藻類,分佈局限於透光帶內。分佈於古新世至現代。
多囊蟲目(Polycystina)
中心囊膜由密集並列的有機質小板構成,小板上具小孔。共生的溝鞭藻位於囊外。骨骼成分為硅質。根據殼體結構和對稱性,分為: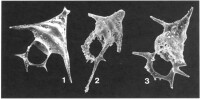 泡沫蟲亞目(Spumellina)、罩籠蟲亞目(Nassellina)、阿爾拜蟲亞目(Albaillellaria)
泡沫蟲亞目(Spumellina)、罩籠蟲亞目(Nassellina)、阿爾拜蟲亞目(Albaillellaria)
放射蟲
泡沫蟲亞目(Spumellina)
單體或群體,中心囊膜上均勻分佈著許多小孔;極少數的屬種無骨骼;少數的屬種具分散於外質中的骨針;大多數泡沫蟲個體微小,具有單一的格子殼——典型形態(球形對稱);還有一些屬種具放射狀刺和內部支持橫桿的同心殼;一些屬種的骨架是由無秩序稠密排列的小棒組成的海綿狀殼。最古老的類群,寒武紀——現代。占放射蟲屬數量的一半。依據骨架的類型和形態以及放射主針數目可劃分出42科。
放射蟲
罩籠蟲亞目(Nassellina)
中心囊膜的穿孔集中在一端,成為一個孔板,構成內囊軸絲圓錐體的基部。殼異極,一般為兩側對稱骨架有主針體型:三射、四射、六射;矢狀環型:簡單或帶刺,具三角形的基角:格架型:由單列的房室構成,分為頂角、頭、胸、腹、基殼口和翼,口孔簡單或被一格架封閉。S出現,古生代屬種較少,中、新生代屬種增加,現代占放射蟲動物群的70%以上。在海洋中分佈廣泛,不同的深度有特定的組合。
阿爾拜蟲亞目(Albaillellaria)
一類已經絕滅的類群,S---P末期。殼體兩側對稱,有一個三角形的骨架支撐,殼壁一般不穿孔。殼體有多種類型
僅具有一封閉的三角形骨架(3);三角形的骨架上部支撐著一個外殼。有的外殼可分為三節,頂錐、假胸和假腹。
褐囊蟲目(Phaeodaria)
由一層稠密的有機質組成。中心囊內有3個孔口,主口側面有褐色色素堆積。大多數骨骼為摻有有機質的蛋白質組成,為分散的刺或格子狀殼。無共生的藻類,常生存於2000米以下的深部水層。化石稀少,白堊紀——現代
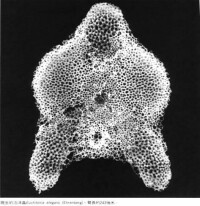
放射蟲
分佈岩性:層狀硅酸岩、深海相沉積岩——黑色頁岩、海相石灰岩、深海相泥岩中的鈣質結核。
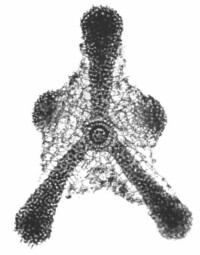
放射蟲
古氣候——放射蟲組分的變化,可推斷相應的溫度波動——丰度、分異度。
恢復古洋流體系——水團
推測古沉積深度(CCD面)——放射蟲軟泥
板塊縫合線附近存在的放射蟲硅質岩的研究具有重要的意義。
根據對不同地層放射蟲種類的發展演化的研究,分析放射蟲的進化譜系,對原生動物的演化有許多啟示。另一方面,由於在世界各大洋深海鑽探獲得大量岩心,對世界大洋洋底地層,自侏羅紀到第四紀各地質時期放射蟲的丰度都有了記錄,許多放射蟲的沉積層序都保存得很好,並且找到了放射蟲在地層中的初現和絕滅時限。因而,利用放射蟲進行生物地層划帶的工作最近有了迅速的發展,其中新生代的划帶工作進展最快。與此同時,基於放射蟲建立的古地磁地層划帶工作,首先在南極地區開展,現在已擴大應用到赤道太平洋和北大西洋兩水域的海底地層。古地磁方法認為放射蟲的絕滅種類與地磁倒轉相應,但對古地磁方法的有效性,學者們仍有不同的意見。此外,不少學者指出放射蟲的分佈與水團的邊界相應,某些放射蟲成為某些水團的地方種,因此,有人認為放射蟲可以作為水團的指示種。至於用放射蟲解釋古氣候、作為古氣候指標,也是近年來對放射蟲應用的發展。放射蟲是海洋環境生活的原生動物,淡水環境絕無生存,因此,它又是海相的指示生物。在大陸架地區,觀察它的數量變化規律,可以探究該地區海進與海退的演替。
1、《微體古生物學教程》,第二版,中國地質大學出版社