共找到12條詞條名為寒武紀的結果 展開
寒武紀
地質年代
寒武紀(Cambrian)是顯生宙的開始,距今約5.42億年前—4.88億年。這個名字來自於英國威爾士的一個古代地名羅馬名稱“Cambria”,該地的寒武紀地層被最早研究。“寒武紀”一詞是英國地質學家塞奇威克(A Sedgwick)於1835年首次引進地質文獻的。原指泥盆紀老紅砂岩之下的所有地層。在羅馬人統治的時代,北威爾士山曾稱寒武山,因此賽德維克便將這個時期稱為寒武紀。通過鈾鉛測年法測量其延續時間為5370萬年。
中文名稱源自舊時日本人使用日語漢字音讀的音譯名“寒武紀”(音讀:カンブキ,羅馬字:kanbuki)。
寒武紀可再分為早寒武紀、中寒武紀、晚寒武紀。寒武紀是顯生宙最早的地質時代,下一個紀是奧陶紀。在傳統理論上它可區分為三個時期:早寒武紀(5.41-5.4億年前)、中寒武紀(5.4-5.23億年前)、以及晚寒武紀(5.23-4.92億年前)。這是建立在岩石地層學上的分法。近幾年國際地層委員會在中國科學家研究的基礎上確立了“四分法”的寒武紀新年代地層表。
2005年之後,國際地層委員會將下寒武統拆分,使得寒武系(即寒武紀時期形成的地層)由原來的三分變為四分,起始年代也有進一步修正。

寒武紀
這一時期,最繁榮的生物是節肢動物三葉蟲,其次是腕足動物、古杯動物、棘皮動物和腹足動物。寒武紀的生物形態奇特,和地球上的現生生物極不相同。最古老的魚種也出現在這個時代,是耳材村海口魚(Haikouichthys ercaicunensis),該化石發掘在澄江動物群。
此時在潮濕的低地可能分佈有苔蘚和地衣類的低等植物,但它們還缺乏真正的根莖組織,難以在乾燥地區生活。寒武紀沒有真正的陸生生物。
寒武紀是最早的地質時代,下一個紀是奧陶紀。
“寒武”源自英國威爾士的古拉丁文“Cambria”。日文音譯,中國沿用。1936年賽德維克在英國西部的威爾士一帶進行研究,在羅馬人統治的時代,北威爾士山曾稱寒武山,因此賽德維克便將這個個時期稱為寒武紀。
寒武紀分為早寒武世、中寒武世和芙蓉世。動物群以具有堅硬外殼的、門類眾多的海生無脊椎動物大量出現為其特點,是生物史上的一次大發展。其中三葉蟲最為常見,是劃分寒武系的重要依據。其它尚有無鉸、幾丁質外殼的腕足類小舌形貝、小園貨貝以及古杯類和軟舌螺等。植物群以藻類為主,還有一些微古植物。
寒武紀三葉蟲群分區現象特別明顯。動物地理區主要有兩個,即東方太平洋區和西方大西洋區。大西洋動物群,其分佈範圍包括大西洋兩岸,如西北歐及美洲最東部新英格蘭地帶。早寒武世、中寒武世和芙蓉世分別以賀爾姆蟲、奇異蟲和油櫛蟲為代表。太平洋動物群,則以萊德利基蟲、庫庭蟲和褶盾蟲為代表。
2005年,國際地層委員會在中國科學家研究的基礎上確立了“四分法”的寒武紀新年代地層表,寒武紀被分為四個世:紐芬蘭世(541.0±1.0-約521Ma)、第二世(約521-約509Ma)、苗嶺世(約509-497Ma)、芙蓉世(497-485.4±1.9Ma)(加約的部分為未確定GSSP【金釘子】)。

身形像豬和有類似豬鼻的水龍獸,據說在恐龍時代來臨前曾統治地球超過100萬年
宙下被劃分為一些代。通常的分法大致有:太古代、元古代、古生代、中生代、新生代五個代。太古代一般指的是地球形成及化學進化這個時期,可以是從46億年前到38億年前或34億年前,這個數字之所以有數以億計的年數之差是因為我們目前所能掌握的最古老的生命或生命痕迹還有許多的不確定因素。元古代緊接在太古代之後,其下限一般定在前寒武紀生命大爆發之前,這個時期目前在5.7億到6億年前。太古代和元古代這兩個名稱是1863由美國人洛岡命名的,他命名的意思是指生物界太古老和生物界次古老。自寒武紀後到2.3億年前這段時間為古生代,這個名稱由英國人賽德維克制定,他依照洛岡取了生物界古老的意思,此事發生在1838年。從2.3億年前到0.65億年前為中生代,從0.65億年後到現在為新生代。這兩個代均由英國人費利普斯於1841年命名,取意分別為生物界中等古老和生物界接近現代。
寒武紀對我們來說是十分遙遠而陌生的,這個時期的地球大陸特徵完全不同於今天。寒武紀常被稱為“三葉蟲的時代”,這是因為寒武紀岩石中保存有比其他類群豐富的礦化的三葉蟲硬殼。當時出現了豐富多樣且比較高級的海生無脊椎動物,保存了大量的化石,從而有可能研究當時生物界的狀況,並能夠利用生物地層學方法來劃分和對比地層,進而研究有機界和無機界比較完整的發展歷史。
地球開始爆發了全球性的加里東構造運動,地球各板塊互相碰撞、擠壓,火山噴發頻繁,南、北大陸發生多次反覆升降,許多地區重新又被海水所淹沒。北方各大陸的相對位置較震旦紀時靠近,但互相併沒有連接;南方岡瓦納大陸逐漸連在一起,但並不是完全相連,且可能通過非洲與北方大陸相連通。寒武紀,最主要的特徵是生物界的顯著繁盛和化石的大量保存。早期,氣候乾旱,淺海分佈廣泛,門類眾多的海洋無脊椎動物突然之間出現在地球上,這是生物史上的第一次大發展,被稱為著名的“生命大爆發”事件。從此,地球上的生物開始了大發展時期,並且空前繁盛,海水裡出現了許許多多種動物,如三葉蟲、頭足類、珊瑚等。
被稱為古生物學和地質學上的一大懸案──寒武紀生命大爆發,自達爾文以來就一直困擾著進化論等學術界。大約6億年前,在地質學上稱做寒武紀的開始,絕大多數無脊椎動物門在幾百萬年的很短時間內出現了。這種幾乎是“同時”地、“突然”地出現在寒武紀地層中門類眾多的無脊椎動物化石(節肢動物、軟體動物、腕足動物和環節動物等),而在寒武紀之前更為古老的地層中長期以來卻找不到動物化石的現象,被古生物學家稱作“寒武紀生命大爆發”,簡稱“寒武爆發”。達爾文在其《物種起源》的著作中提到了這一事實,並大感迷惑。他認為這一事實會被用做反對其進化論的有力證據。但他同時解釋到,寒武紀的動物一定是來自前寒武紀動物的祖先,是經過很長時間的進化過程產生的;寒武紀動物化石出現的“突然性”和前寒武紀動物化石的缺乏,是由於地質記錄的不完全或是由於老地層淹沒在海洋中的緣故。
寒武紀生命大爆發的起源—埃迪卡拉動物群
埃迪卡拉(Ediacaran)動物群是Sprigg於1947年在澳大利亞中南部Ediacara地區的龐德砂岩層中首先發現的。最初人們未能確定這一動物群的時代,後來終於確定為前寒武紀,年齡為6.7億年。埃迪卡拉動物群包含三個門,19個屬,24種低等無脊椎動物。三個門是:腔腸動物門,環節動物門和節肢動物門。水母有7屬9種;水螅綱有3屬3種;海鰓目(珊瑚綱)有3屬3種;缽水母2屬2種;多毛類環蟲2屬5種;節肢動物2屬2種。多保存為印痕化石,儘管它們的形態、結構都很原始,但它們被認為是20世紀古生物學最重大的發現之一。這一發現使科學界擯棄了長期以來認為在寒武紀之前不可能出現後生動物化石的傳統觀念。所謂後生動物即是指相對於原生動物的各種多細胞動物。
艾迪卡拉動物群包含了多種形態奇特的動物化石:身體巨大而扁平、多呈橢圓形或條帶形,具有平滑的有機質膜,是人們迄今為止發現的最古老、最原始的化石,也是在太古代地層中發現的最有說服力的生物證據。按Seilacher的觀點,艾迪卡拉動物群可分為輻射狀生長、兩極生長和單極生長3種類型。除輻射狀生長的類型中可能有與腔腸動物有關係的類群外,其他兩類與寒武紀以後出現的生物門類無親源關係。
儘管有關艾迪卡拉(型)動物群的性質還有許多爭議,但其奇怪的形態令許多學者相信,艾迪卡拉(型)動物群是後生動物出現后的第一次適應輻射,它們採取的不同於現代大多數動物採取的形體結構變化方式。不增加內部結構的複雜性,只改變軀體的基本形態,變得非常薄,成條帶狀或薄餅狀,使體內各部分充分接近外表面,在沒有內部器官的情況下進行呼吸和攝取營養。如現代大型寄生動物滌蟲現代大多數動物採取的是保持渾圓或球形的外部形態的同時,進化出複雜的內部器官來擴大相應的表面積(如肺、消化道),從化石上可以看出,這些生物已具有了高度分化的組織和器官,說明它們已不是最原始的類型。它們代表了後生動物出現以後的第一次輻射演化因此,可以認為艾迪卡拉(型)動物群是在元古宙末期大氣氧含量較低的條件下後生動物大規模佔領淺海的一次嘗試,結果失敗了,而導致絕滅。在後來的演化過程中,後生動物採取了第二種方式,使內部的器官複雜化和物種多樣化的發展,即生物系統演化。
寒武紀生命大爆發的代表─雲南澄江動物群
寒武爆發的典型代表是被稱為20世紀最驚人的科學發現之一的中國雲南澄江動物群,它是世界上目前所發現的最古老、保存最為完整的帶殼後生動物群。該動物群是中國青年古生物學家侯先光1984年在雲南澄江縣帽天山首先發現的。這是一個內容十分豐富、保存非常完美,距今約5.7億年的化石群,其成員包括水母狀生物、三葉蟲、具附肢的非三葉的節肢動物、金臂蟲、蠕形動物、海綿動物、內肛動物、環節動物、無絞綱腕足動物、軟舌螺類、開腔骨類,以及藻類等,甚至還有屬於低等脊索動物或半索動物(如著名的雲南蟲)等。由於許多動物的軟組織保存完好,為研究早期無脊椎動物的形態結構、生活方式、生態環境等提供了極好的材料,同時也成為了探索地球上大殼後生動物爆發事件的重要窗口。

寒武紀海洋生態系統中最頂級的捕食者“奇蝦”
1965年,兩位美國物理學家提出了寒武爆發是由於地球大氣的氧水平這個物理因素造成的。
他們認為,在早期地球的大氣中含有很少或根本就沒有自由氧,氧是前寒武紀藻類植物光合作用的產物並逐漸積累形成的。後生動物需要大量的氧,一方面用於呼吸作用,另一方面氧還以臭氧的形式在大氣中吸收大量有害的紫外線,使後生動物免於有害輻射的損傷。
生物學家則從生物本身的生態關係來探討這一問題,因為地質學的證據否定了這種氧理論的觀點。大約在距今10億年至20億年之間廣泛沉積層中含有大量嚴重氧化的岩石,這說明在這一時期內已經存在足夠生命爆發的氧條件。因而生物學家從兩個重要事件的出現來探索造成寒武爆發的原因,即有性生殖的產生和生物收割者的出現。
從化石資料來看,真核藻類大約在9億年前出現了有性生殖,實際上,有性生殖出現得更早。有性生殖的發生在整個生物界的進化過程中有著極其重大的作用,由於有性生殖提供了遺傳變異性,從而有可能進一步增加了生物的多樣性,這是造成寒武爆發的原因之一。
生物收割者假說是美國生態學家斯坦利提出的,是一種解釋寒武爆發的生態學理論,即收割原則。斯坦利認為,在前寒武紀的25億年的多數時間裡,海洋是一個以原核藍藻這樣簡單的初級生產者所組成的生態系統。這一系統內的群落在生態學上屬於單一不變的群落,營養級也是簡單唯一的。由於物理空間被這種種類少但數量大的生物群落頑強地佔據著,所以這種群落的進化非常緩慢,從未有過豐富的多樣性。寒武爆發的關鍵是草食收割者的出現和進化,即食用原核細胞(藍藻)的原生動物的出現和進化。收割者為生產者有更大的多樣性製造了空間,而這種生產者多樣性的增加又導致了更特異的收割者的進化。營養級金字塔按兩個方向迅速發展:較底層次的生產者增加了許多新物種,豐富了物種多樣性,在頂端又增加了新的“收割者”,豐富了營養級的多樣性。從而使得整個生態系統的生物多樣性不斷豐富,最終導致了寒武紀生命大爆發的產生。
對於“收割理論”科學家們目前還沒有找到直接的證據來證明其正確性,然而,一些間接的證據支持了這一理論。間接證據之一來自於前寒武紀疊層石,這些由藻類組成的疊層石中保存了前寒武紀最豐富的生產者群落。今天,疊層石僅盛產於缺少後生動物收割者的貧瘠環境中,如超鹽量的鹹水湖中。藻類在前寒武紀地層中的大量存在,大概反映了當時收割者的貧乏。另外,生態學野外研究也提供了一些間接的證據,研究表明,在一個人工池塘中,放進捕食性魚,會增加浮游生物的多樣性;從多樣的藻類群落中去掉海膽,會使某一藻類在該群落中占統治地位而多樣性下降。
寒武爆發作為地史上的第二大懸案一直為人們所關注。隨著化石的不斷發現及新理論的建立,這一謎團最終將大白於天下。
在寒武世時,具有硬殼的動物第一次大量地出現,許多大陸都被淺海所泛濫,超大陸剛瓦那(Gondwana)則正在南極附近形成。
形成於前寒武世末期(大約距今六億年前)的超大陸潘諾西亞(Pannotia),在古生代的時候開始四分五裂,一個新的海洋--巨神海(Iapetus Ocean)在勞倫西亞(Laurentia,北美)、波羅地(Baltica,北歐)和西伯利亞(Siberia)這幾個古大陸之間擴張。超大陸岡瓦那(Gondwana)則在泛非褶皺帶上組合而成當時最大的大陸,範圍從赤道延伸到南極。
在奧陶世時期,溫暖的海水把石灰岩和鹽岩沉澱在岡瓦那大陸的赤道地區(Australia澳洲、India印度、China中國與Antarctica南極洲),同時在岡瓦那大陸的南極地區(Africa非洲與South America南美)則沉澱了冰河的沉積及冰漂的碎屑。
也稱“前古生代”,古生代第一個紀-寒武紀(距今約六億年)之前的地質時代。
寒武紀的開始,標誌著地球進入了生物大繁榮的新階段。而在寒武紀之前,地球早已經形成了,只是在幾十億年的漫長過程中一片死寂,那時地球上還沒有出現門類眾多的生物。這樣,科學家們便把寒武紀之前這一段漫長而缺少生命的時間稱作前寒武紀。前寒武紀約佔全部地史時間的六分之五,根據有關生命活動跡象的寶貴資料,也是為了研究上的便利,地質學家把漫長的前寒武紀分為太古代、元古代兩部分。
太古代之前(地球形成之初-38億年前)則有多種不同的稱呼。太古代離我們久遠,其時限約從38億年至26億年前,長達12億年。太古代是具有明確地史記錄的最初階段。在這漫長的12億年間,是地球形成后的初始期,地表到處形成童山和荒漠,由於年代久遠,確實很難尋覓到化石,人們對這一時期的生命活動了解得很少。但20世紀後半期,科學家們陸續在南非和澳大利亞獲得了重大收穫,在變質程度不太劇烈的沉積岩層中發現了疊層石,這是微生物和藻類活動的產物。此外,人們在這些古老的岩層中還分析出大量的有機化合物(如苯、烴基苯等)和環形化合物(如呋喃、甲醇、乙醛等)。在南非的一套古老沉積岩中,科學家們藉助先進的精密觀測儀器,發現了200多個與原核藻類非常相似的古細胞化石,這些微體化石一般為橢圓形,具有平滑的有機質膜,這是人們迄今為止發現的最古老、最原始的化石,也是在太古代地層中發現的最有說服力的生物證據。從生物界看,這是原始生命出現及生物演化的初級階段,當時只有數量不多的原核生物,他們只留下了極少的化石記錄。從非生物界看,太古宙是一個地殼薄、地熱梯度陡、火山—岩漿活動強烈而頻繁、岩層普遍遭受變形與變質、大氣圈與水圈都缺少自由氧、形成一系列特殊沉積物的時期;也是一個硅鋁質地殼形成並不斷增長的時期,又是一個重要的成礦時期。
元古代的時限自26億年前至5.7億年,在這段地史中,原核生物演化為真核細胞生物,形成地史時期的菌-藻類時代。人們在這一時期的古老地層中發現過微古植物化石、宏觀藻類化石及疊層石。僅在我們中國,古生物學家就已發現元古代不同時期的微古植物化石80餘屬、近200個種,生命在元古代得到進一步繁榮,那時的地球已不再是滿目荒蕪了。初期地表已出現了一些範圍較廣、厚度較大、相對穩定的大陸板塊。因此,在岩石圈構造方面元古代比太古代顯示了較為穩定的特點。早元古代晚期的大氣圈已含有自由氧,而且隨著植物的日益繁盛與光合作用的不斷加強,大氣圈的含氧量繼續增加。元古代的中晚期藻類植物已十分繁盛,明顯區別於太古代。

昆蟲遠祖的遠祖----撫仙湖蟲生態景觀
之後美國古生物學家畢可脫(1913年)、日本人遠藤隆次(1939年)、小林貞一(1941年、1942年、1955年)均相繼研究過張夏、崮山一帶寒武紀地層中的生物化石。我國著名的地質學家孫雲鑄教授從1923年起對張夏、崮山地區的寒武系進行了長達20餘年的研究,對寒武紀地層做了劃分。1953年盧衍豪、董南庭重新觀察了張夏、崮山一帶寒武紀標準剖面,其中最重要的是把B·維里斯和E·布萊克威爾德的饅頭頁岩自下而上再分為饅頭組、毛庄組、徐庄組,並把前兩個組置於下寒武統,把后一個組歸入中寒武紀,炒米店灰岩再分為鳳山組及長山組,將張夏地區寒武系確定為7個單位和17個三葉蟲化石帶。此後,有關地質院校,如北京地質學院等,以及山東省地質局等單位,先後對張夏寒武紀地層剖面作過大量的野外觀察、剖面測制、室內鑒定和專題研究,取得了豐富的實際資料,從不同的角度補充和完善了該剖面的基礎資料,進一步提高了該剖面的研究水平。
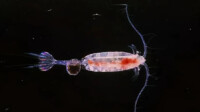
浮遊雙瓣殼節肢動物
1953年盧衍豪、董南庭重新觀察了張夏、崮山一帶寒武紀標準剖面,其中最重要的是把B·維里斯和E·布萊克威爾德的饅頭頁岩自下而上再分為饅頭組、毛庄組、徐庄組,並把前兩個組置於下寒武統,把后一個組歸入中寒武統,炒米店灰岩再分為鳳山組及長山組,將張夏地區寒武系確定為7個單位和17個三葉蟲化石帶。此後,有關地質院校,如北京地質學院等,以及山東省地質局等單位,先後對張夏寒武紀地層剖面作過大量的野外觀察、剖面測制、室內鑒定和專題研究,取得了豐富的實際資料,從不同的角度補充和完善了該剖面的基礎資料,進一步提高了該剖面的研究水平。
1959年,位於泰山北側的張夏寒武紀地層剖面在全國地層會議上被正式定為華北寒武系標準剖面。1958-1961年,北京地質學院在泰山地區進行1:20萬區調,將泰山變質岩命名為太古代泰山群,自下而上劃分為萬山莊、雁翎關、山草峪組等。1960-1962年,山東地質局805隊開展包括泰安南留幅等23幅1∶5萬區調聯測。1963-1965年,山東地質局805隊進行1∶5萬泰安幅區調,將泰山變質岩稱為泰山雜岩,劃分為望府山、笤帚峪、唐家莊、盂家莊、馮家峪等五個岩組。同時,地科院程裕淇等,山東地質局805隊鄭良峙、張成基等人開展變質岩專題研究,確立了雁翎關、山草峪組層序,恢復了原岩,進行變質岩的岩石學研究。
1978年以後,泰山的地質研究工作,進入了一個以專題研究為主的深入發展級段。泰山的保護、管理和開發建設得到了很大的加強,泰山的面貌有了巨大的變化。1980年,應思淮研究員對泰山變質岩進行了專題研究,並出版專著《泰山雜岩》。1981-1984年,山東礦業學院呂朋菊教授對泰山的形成進行了專題研究,並發表了《泰山形成及其年齡》一文。1982-1985年,山東區調隊鄭良峙、王世進等人進行了魯西泰山群專題研究,新建柳杭組,置於雁翎關組和山草峪組之上。1982-1987年,山東地礦局第一地質隊馬雲順、翟穎川等對魯西太古代綠岩帶含礦性進行專題研究。1982-1984年,趙世英等進行紅門“桶狀構造”的專題研究,並發表《泰山紅門“桶狀構造”成因的探討》論文。1982-1985年,地科院朱振華碩士填制了泰山山前1∶2.5萬地質圖,完成碩士論文,發表“泰山太古宙岩漿雜岩體的岩石化學和地球化學特徵”論文。
1983-1986年,法國雷恩大學江博明等,地科院沈其韓等,山東地礦局董一傑等對中國太古宙地殼演化進行專題研究,認為泰山雜岩的大部分為變質侵入岩(灰色片麻岩),稱之為望府山片麻岩,取得大量地球化學和同位素年齡資料。1986-1987年,北京大學謝凝高等進行泰山風景名勝區資源綜合考察評價及其保護利用研究。1986-1990年,山東區調隊王世進等進行1∶20萬泰安、新泰幅修測。同期,地科院徐惠芬、山東地質一大隊董一傑等人對山東魯西太古宙綠岩帶和魯西太古代地層等進行專題研究,系統總結了泰山岩群的分佈、層序、變質作用的特點,並出版專著。
1987年聯合國教科文組織把泰山列為世界自然與文化遺產,泰山的地位和影響發生了歷史性的變化。1989-1990年,山東礦業學院呂朋菊教授等對泰山周圍重力滑動構造進行專題研究,發表了“泰山周圍太古界與古生界不整合面上滑動構造的發現”論文。1990年,山東省地礦局第一地質大隊董一傑等發表《泰山地區太古宙雜岩體的地球化學特徵》論文。1990-1993年,山東地礦局曹國權等人發表《魯西早前寒武紀地質》專著。1993-1996年,地質礦產部地質研究所、山東第一地質礦產勘查院進行泰安市幅1∶5萬區域地質調查。
1994-1995年,呂朋菊教授就泰山新構造運動的進行專題研究,發表了《新構造運動與現今泰山的形成及地貌景觀》論文,並對泰山地質地貌進行總結,撰寫《泰山大全》之地質地貌篇。1995年地科院庄育勛等,山東地礦局第一地質大隊張富中等就泰山地區新太古代~古元古代地殼演化研究的新進展發表論文。1997年,地科院庄育勛等,山東地礦局第一地質大隊任志康等人在《岩石學報》發表論文《泰山地區早前寒武紀主要地質事件與地殼演化》。1998年,山東地勘局地質調查研究院呂發堂等就其研究成果發表《泰山地區晚太古代“框架侵入岩”的地質特徵及稀土地球化學演化》論文。
1999年,地科院王新社等,山東地礦局第一地質大隊任志康等發表《泰山地區太古宙末韌性剪切作用在陸殼演化中的意義》論文。2000年,地科院地質力學所張明利等發表《新生代構造運動與泰山形成》論文。2002-2003年,山東科技大學呂朋菊教授等、泰山風景區管委會牛健等人進行泰山地質地貌特徵及地學價值評價專題研究,並發表《泰山的地學價值及其意義》論文。2004年9月中國地質大學對擬建中國泰山世界地質公園的建設條件、地質遺跡和資源狀況等進行了為期兩周的研究考察。
張夏寒武紀地層剖面,把寒武系劃分為下、中、上統的7個地層單位,即下統的饅頭組,中統的毛庄組、徐庄組、張夏組,上統的崮山組、長山組、鳳山組。現從老到新簡述如下:饅頭組 主要由紫紅色、黃綠色等雜色頁岩及泥質、白雲質灰岩組成。底部不整合於泰山雜岩的肉紅色片麻狀花崗岩之上。下部灰岩中含磁石結核和條帶,上部頁岩中具微細水平層理,中部頁岩含有三葉蟲化石~ 中華萊德利基蟲。厚度119米。
毛庄組 主要由紫色雲母質頁岩和灰岩組成。含三葉蟲、腕足類及藻類化石。厚度39米。
徐庄組 主要由紫灰色頁岩和鮞狀灰岩組成,其中下部的灰岩及灰質粉砂岩中常具斜層理或交錯層理。含有徐庄蟲等三葉蟲化石及腕足類化石。厚度73米。
張夏組 主要由鮞狀灰岩和藻類灰岩組成,中夾雜色頁岩。含德氏蟲等三葉蟲化石。厚度198米。
崮山組 主要由竹葉狀灰岩、疙瘩狀灰岩和黃綠色頁岩組成。含蝴蝶蟲、蝙蝠蟲等三葉蟲化石。厚度51米。
長山組 主要由迭層石灰岩、具紅色氧化圈竹葉狀灰岩、紫色頁岩組成。含有庄氐蟲、蒿里山蟲等三葉蟲化石。厚度70米。
鳳山組 主要由泥質灰岩和竹葉狀灰岩組成。含濟南蟲、方頭蟲等三葉蟲化石,以及海百合莖和腕足類化石。厚度130米。
張夏寒武紀地層的標準剖面,分別位於張夏和崮山一帶的饅頭山、虎頭崖、黃草頂、唐王寨、范庄等地。其中的饅頭山是徐庄組、毛庄組、饅頭組的剖面,虎頭崖~黃草頂是張夏組的剖面,唐王寨是崮山組、長山組的剖面,范庄是鳳山組的剖面。
饅頭山位於張夏鎮南2公里,104國道的西側。因其狀如饅頭而得名。山的北麓有兩個名為徐庄和毛庄的小村子。它是張夏寒武紀標準地層剖面的饅頭組、毛庄組、徐庄組的建組和命保所在地。
張夏寒武紀地層剖面,在泰山主峰之北,位於泰安和濟南之間交通幹線的兩側,交通方便,而且構造簡單,出露完全,十分有利於現場觀察和研究。它是我國地層和古生物研究歷史最長、研究程度最高的地層剖面之一,在我國地質學史上佔有很重要地位。1959全國地層會議后,被正式確定為我國北方寒武系的標準地層剖面,在我國不同地區寒武紀地層對比和國際寒武紀地層對比方面起著重要作用,同時也是許多寒武紀古生物種屬(蒿里山蟲Kaolizhania、中華萊德利基蟲 Redichia Chinensis、饅頭褶頰蟲Ptychopariamantoensis、山東蟲Shantungia、孫氏盾蟲Sunasp-is Lavevis)命名地或模式標本的原產地。因此,這個標準地層剖面,在國內外十分聞名,長期以來有許多國內外地質學者不斷來此參觀考察,同時也成為我國大專院校地學的重要實習基地,無論在地質科學方面,還是在生產實踐以及地質教育方面,都具有很高的科學價值
寒武紀是地質年代劃分中屬顯生宙古生代的第一個紀,距今約5.7億至5.1億年,寒武紀是現代生物的開始階段,是地球上現代生命開始出現、發展的時期。寒武紀對我們來說是十分遙遠而陌生的,這個時期的地球大陸特徵完全不同於今天。寒武紀常被稱為“三葉蟲的時代”,這是因為寒武紀岩石中保存有比其他類群豐富的礦化的三葉蟲硬殼。
此外,寒武紀還產生了進化史上的一個重要事件“寒武紀生命大爆發”,在很短(地質意義上的很短,其實也有數百萬年之久)時間內,生物種類突然豐富起來,呈爆炸式的增加。它意味著,生物進化除了緩慢漸變,還可能以跳躍的方式進行。當時出現了豐富多樣且比較高級的海生無脊椎動物,保存了大量的化石,從而有可能研究當時生物界的狀況,並能夠利用生物地層學方法來劃分和對比地層,進而研究有機界和無機界比較完整的發展歷史。但澄江生物群告訴我們,如今地球上生活的多種多樣的動物門類在寒武紀開始不久就幾乎同時出現。
寒武紀是顯生宙(Phanerozoic Eon)的開始,標誌著地球生物演化史新的一幕。在寒武紀開始后的短短數百萬年時間裡,包括現生動物幾乎所有類群祖先在內的大量多細胞生物突然出現,這一爆髮式的生物演化事件被稱為“寒武紀生命大爆炸”(Cambrian explosion)。帶殼、具骨骼的海洋無脊椎動物趨向繁榮,它們營底棲生活,以微小的海藻和有機質顆粒為食物,其中,最繁盛的是節肢動物三葉蟲,故寒武紀又稱為“三葉蟲時代”,其次是腕足動物、古杯動物、棘皮動物和腹足動物,寒武紀的生物形態奇特,和我們現在地球上所能看見的生物極不相同。比較著名的有早寒武世雲南的澄江生物群、加拿大中寒武世的布爾吉斯頁岩生物群。寒武紀的生物界以海生無脊椎動物和海生藻類為主。無脊椎動物的許多高級門類如節肢動物、棘皮動物、軟體動物、腕足動物、筆石動物等都有了代表。其中以節肢動物門中的三葉蟲綱最為重要,其次為腕足動物。此外,古杯類、古介形類、軟舌螺類、牙形刺、鸚鵡螺類等也相當重要。拋開牙形石不說,高等的脊索動物還有許多其他代表,如中國雲南澄江生物群中的華夏鰻、雲南魚、海口魚等,加拿大布爾吉斯頁岩中的皮開蟲,美國上寒武紀的鴨鱗魚。
在潮濕的低地,可能分佈有苔蘚和地衣類的低等植物,但它們還缺乏真正的根莖組織,難以在乾燥地區生活;無脊椎動物也還沒有演化出適應在空氣中生活的機能。寒武紀沒有真正的陸生生物,大陸上缺乏生氣、荒涼一片。
古生物學引用“大爆發”一詞,來形容生物多樣性突然爆髮式出現。根據寒武紀開始時痕迹化石和小殼化石的突然多樣性和複雜性,“寒武紀大爆發”的理論在澄江生物群發現之前就已提出,但對“寒武紀大爆發”所產生的動物及動物群落結構特徵所知甚微,即使著名的加拿大布爾吉斯頁岩動物群化石也比“寒武紀大爆發”晚1000多萬年,不能回答寒武紀初期海洋中具體有什麼生命。
澄江生物群的地質時代正處於“寒武紀大爆發”時期,它讓我們如實看到5.3億年前動物群的真實面貌,各種各樣的動物在“寒武紀大爆發”時期迅速起源,立即出現,它將動物多樣性的歷史前推到寒武紀早期。
寒武紀的生物形態奇特,和地球上的現生生物極不相同。最古老的魚種也出現在這個時代,是耳材村海口魚(Haikouichthys ercaicunensis),該化石發掘在澄江生物群。
在傳統的岩石地層學上將寒武紀三分為早寒武世、中寒武世和晚寒武世。
但是根據國際地層學研究的新要求,自2005年起國際上使用了新的年代地層學的寒武紀四分法來取代三分法,具體劃分如下:
| 世 | 期 | 期起始時間(Ma) | GSSP | 標誌事件 |
| 幸運世 | 第十期 | 489.5 | (候選)中國浙江江山縣碓邊村剖面 | 三葉蟲類Lotagnostus americanus首現,顯微鏡下的觀察表明牙形石Codylodus adesei 可能首現 |
| 江山期 | 494 | 中國浙江江山縣碓邊B剖面 | 球接子類三葉蟲Agnostotes orientalis首現,polymerid trilobite類的Irvingella angustilimbata 消失 | |
| 排碧期 | 497 | 中國湖南花垣縣武陵山 | 三葉蟲類Glyptagnostus reticulatus首現 | |
| 苗嶺世 | 古丈期 | 500.5 | 中國湖南古丈縣羅依溪 | 三葉蟲類Lejopyge laevigata首現 |
| 鼓山期 | 504.5 | 美國猶他州Millard縣鼓山 | 三葉蟲類Ptychagnostus atavus 首現 | |
| 烏溜期 | 509 | 中國貴州烏溜-曾家岩剖面 | 三葉蟲類 Oryctocephalus indicus首現 | |
| 第二世 | 第四期 | 514 | (尚無) | 小油櫛蟲屬(Olenellus)或萊德利基蟲屬(Redlichia)首現 |
| 第三期 | 521 | (尚無) | 三葉蟲類首現 | |
| 紐芬蘭世 | 第二期 | 529 | (尚無) | 小殼化石,或古杯動物 |
| 幸運期 | 541.0±1.0 | 加拿大紐芬蘭東南部Fortune Head | 遺跡化石Trichiophycus pedum 首現 |
與我們一般的認識不同,作為寒武紀生命大爆發的代表的三葉蟲,是在第二統的早期才出現的,而不是寒武紀的一開始。
寒武紀動物群以具有堅硬外殼的、門類眾多的海生無脊椎動物大量出現為其特點,是生物史上的一次大發展。其中三葉蟲最為常見,是劃分寒武系的重要依據。其它尚有無鉸、幾丁質外殼的腕足類小舌形貝、小園貨貝以及古杯類和軟舌螺等。植物群以藻類為主,還有一些微古植物。寒武紀三葉蟲群分區現象特別明顯。
動物地理區主要有兩個,即東方太平洋區和西方大西洋區。大西洋動物群,其分佈範圍包括大西洋兩岸,如西北歐及美洲最東部新英格蘭地帶。早寒武世、中寒武世和晚寒武世分別以賀爾姆蟲、奇異蟲和油櫛蟲為代表。太平洋動物群,則以萊德利基蟲、庫庭蟲和褶盾蟲為代表。
寒武紀生命大爆發被稱為古生物學和地質學上的一大懸案。寒武紀生命大爆發自達爾文以來就一直困擾著進化論等學術界。大約6億年前,在地質學上稱做寒武紀的開始,絕大多數無脊椎動物門在只2000多萬年時間內出現了。這種幾乎是“同時”地、“突然”地在2000多萬年時間內出現在寒武紀地層中門類眾多的無脊椎動物化石(節肢動物、軟體動物、腕足動物和環節動物等),而在寒武紀之前更為古老的地層中長期以來卻找不到動物化石的現象,被古生物學家稱作“寒武紀生命大爆發”,簡稱“寒武爆發”。
達爾文在其《物種起源》的著作中提到了這一事實,並大感迷惑。他認為這一事實會被用做反對其進化論的有力證據。但他同時解釋到,寒武紀的動物的祖先一定是來自前寒武紀動物,是經過很長時間的進化過程產生的;寒武紀動物化石出現的“突然性”和前寒武紀動物化石的缺乏,是由於地質記錄的不完全或是由於老地層淹沒在海洋中的緣故。這就是至今仍被國際學術界列為“十大科學難題”之一的“寒武紀生命大爆發”。
依照傳統和經典的生物學理論,即達爾文生物進化認為,生物進化經歷了從水生到陸地、從簡單到複雜、從低級到高級的漫長的演變過程,這一過程是通過自然選擇和遺傳變異兩個車輪的緩慢滾動逐漸實現的。科學家們為揭示“寒武紀大爆發”的原因提出了種種假說。陳均遠研究員提出寒武紀生物突變具有極明顯的自發性進化行為的設想;舒德干教授提出“寒武紀暖水與冷水兩大古生物地理分區"的假說等,但目前還沒有一個清晰、證據確鑿、令人信服的解釋。
地球膨裂說認為“雪球地球’冰期后地溫急聚升高是寒武紀生命大爆發的真正原因。“雪球地球”理論認為,地球在距今7.5億到5.8億年前曾經經歷了一次極其嚴重而漫長的冰河時代——瓦蘭吉爾期。當時不僅陸地全部被冰川覆蓋,海洋表面也被完全凍結,液態水靠來自地球核心的熱量支持,存在於1公里厚的冰層下。如果從太空看,地球完全是一巨大的“雪球”。“雪球地球”假說最早由美國地質學家約瑟夫·可西文克博士1992年首先提出。
“雪球地球”的假說有哪些根據?第一,地球在距今6億到8億年間廣泛發育了一層或多層稱為“冰積岩”的冰川沉積,它代表了全球性的寒冷氣候。最著名的一次冰期發生在距今6億年左右,幾乎在現今所有大陸上都留下了可靠的記錄,地質學上稱為瓦倫格冰期。不管怎樣,在地球上很好保存了距今6億到8億年的地層中,幾乎都能找到同期的冰川沉積。很顯然,這個寒冷氣候是一個全球性事件。第二,在地球的歷史中,很多數據表明,地球在距今6到8億年間,冰積岩大多沉積在中、低緯度附近,換句話說,也就是赤道和赤道附近,也是陸地主要分佈的區域。這方面的資料得出這個結論:廣泛的寒冷氣候發生在地球的赤道及其附近區域。
地球膨裂說認為,雖然生命在38億年前形成,但因為地球逐漸變冷,直到8億年前的“雪球地球”時期的零下50度,物種的進化速度非常緩慢。“雪球”為什麼解凍,變成現在這樣的地球,地球膨裂說認為,8億年前,由於地球內部的放射性物質不斷衰變放出熱量,使地球發生較大膨裂,造成大量岩漿噴出地殼,使地球的氣溫急聚升高,冰川溶化,冰臼就是在這一時期形成的。
地球氣溫從震旦紀(8億年前)的攝氏零下50度,上升到現在的最高溫度攝氏58度,8億年上升了108度,地球的氣溫平均每1千萬年自然升高1.35攝氏度。地球膨裂說認為,8億年前地球的氣溫急聚升高,非常適合生物繁衍,進化速度急聚增加,所以發生了寒武紀生命大爆發。

基本信息
- 中文名
- 寒武紀
- 外文名
- Cambrian
- 年代
- 早寒武紀、中寒武紀、晚寒武紀
- 時間
- 約541±1~485.4±1.9百萬年前
- 主要生物
- 無脊椎動物
- 名稱來源
- 英國威爾士
- 羅馬字
- kanbuki
- 宙
- 顯生宙
- 代
- 古生代
- 劃分
- 四分(幸運世、苗嶺世、第二世、紐芬蘭世)