穿梭系統
穿梭系統
線粒體穿梭(英語:mitochondrial shuttles)是用來轉運還原劑穿過線粒體內膜的體系。還原型煙醯胺腺嘌呤二核苷酸無法穿過該膜,但它可以還原其他可以穿過該膜物質,以便於它所攜帶的電子送達電子傳遞鏈。
在人類中,甘油磷酸穿梭最初被發現於棕色脂肪組織中,正因為這種轉換是低效的,因此褐色脂肪的主要目的就是產生熱量。這種脂肪首先被發現於嬰兒之中,儘管在成人的腎臟周圍和後頸處也存在有少量。蘋果酸-天冬氨酸穿梭被發現於人體的其他部位。
呼吸鏈、生物氧化與氧化磷酸化都是在線粒體內進行的。
線粒體的主要功能是氧化供能,相當於細胞的發電廠。由於線粒體具有雙層膜的結構,外膜的通透性較大,內膜卻有著較嚴格的選擇通透性,通常通過外膜與細胞漿進行物質交換。
已知糖酵解作用是在胞漿(cytosol)中進行的,在真核生物胞液中的NADH不能通過正常的線粒體內膜,要使糖酵解所產生的NADH 進入呼吸鏈氧化生成ATP,必須通過較為複雜的過程,據現在了解,線粒體外的NADH 可將其所帶的H 轉交給某種能透過線粒體內膜的化合物,進入線粒體內后再氧化。即NADH 上的氫與電子可以通過一個所謂穿梭系統的間接途徑進入電子傳遞鏈。能完成這種穿梭任務的化合物有甘油-α-磷酸和蘋果酸等。
在動物細胞內有兩個穿梭系統:磷酸甘油穿梭系統和蘋果酸穿梭系統。但兩種穿梭系統最終等價產能卻不同。
磷酸甘油穿梭(glycerol phosphate shuttle)系統
主要存在於昆蟲的飛翔肌中,靠這一途徑維持很高速度的氧化磷酸化。該途徑主要出現在昆蟲飛行肌中,雖然大多數哺乳動物有此系統,但所佔比重較小。
甘油磷酸穿梭途徑涉及兩個酶:一個是依賴於NAD+的胞液中的甘油-3-磷酸脫氫酶,另一個是嵌線粒體內膜的甘油-3-磷酸脫氫酶複合物,該複合物含有一個FAD輔基和一個位於線粒體內膜外表面的底物結合部位。在胞液甘油-3-磷酸脫氫酶催化下,首先NADH使磷酸二烴丙酮還原生成甘油-3-磷酸,然後甘油-3-磷酸被嵌膜的甘油-3-磷酸脫氫酶複合物(glycerol-3-phosphate dehydrogenase complex)轉換回磷酸二烴丙酮。在轉換過程中兩個電子被轉移到嵌膜酶的FAD輔基上生成FADH2。FADH2將兩個電子轉給可移動的電子載體Q,然後再轉給Q-細胞色素c氧化還原酶,進入電子傳遞鏈。
從總體來看,甘油磷酸穿梭途徑使細胞中的NADH氧化,並在線粒體內膜中生成QH2。胞液中的NADH通過這一途徑轉換成QH2后氧化所產生的能量(1.5個ATP)比線粒體內NADH氧化能量(2.5個ATP)少。
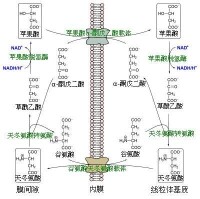
蘋果酸-天冬氨酸穿梭
位於蘋果酸-天冬氨酸穿梭體系中的第一個酶是蘋果酸脫氫酶。蘋果酸脫氫酶在該穿梭體系中有兩種存在形式:線粒體蘋果酸脫氫酶以及胞漿脫氫酶。兩種蘋果酸脫氫酶的區別在於他們的存在位置以及結構,並且在此過程中催化的反應方向相反。
一旦蘋果酸形成,第一個反向轉運體(蘋果酸-α-酮戊二酸)將蘋果酸從胞漿引入線粒體基質與此同時並將α-酮戊二酸從線粒體基質中導出到胞漿中。當蘋果酸到達線粒體基質后,它被線粒體蘋果酸脫氫酶轉換成草醯乙酸,與此同時NAD+被其中的兩個電子還原成NADH且氫離子被釋放出來。草醯乙酸接下來被線粒體天冬氨酸氨基轉移酶轉換為天冬氨酸(因為草醯乙酸不能透過內膜進入胞漿)。因為天冬氨酸是一種氨基酸,為生成它,氨基需要被加到草醯乙酸上。這個氨基由谷氨酸提供,與此同時後者也被同一個酶轉變成了α-酮戊二酸。
第二個反向轉運體(谷氨酸-天冬氨酸)將谷氨酸從胞漿引入線粒體基質與此同時將天冬氨酸從線粒體基質中導出到胞漿中。一旦進入胞漿,天冬氨酸被胞漿天冬氨酸氨基轉移酶轉變成草醯乙酸。
蘋果酸-天冬氨酸穿梭的凈效應是完全地還原:胞漿中的NADH被氧化成NAD+並且線粒體基質中的NAD+被還原成NADH。胞漿中的NAD+接下來可以被另一輪糖酵解還原,而線粒體基質中的NADH可以被用於向電子傳遞鏈傳遞電子以使ATP合成。
因為蘋果酸-天冬氨酸穿梭時線粒體基質中的NADH重新生成,它可以使糖酵解所產生的能量最大化合成ATP(2.5個/NADH),最終導致每個葡萄糖代謝凈收到32個ATP分子。將此與甘油磷酸穿梭相比,後者只將電子傳送給電子傳遞鏈中的複合體II(與還原型黃素腺嘌呤二核苷酸所走路線相同),這樣只能使糖酵解中產生的每個NADH合成1.5個ATP(最終導致每個葡萄糖代謝凈收到30個ATP分子) 。