連鎖遺傳
連鎖遺傳
一種生物的性狀很多,控制這些性狀的基因自然也很多,而各種生物的染色體數目有限,必然有許多基因位於同一染色體上,這就會引起連鎖遺傳的問題。
連鎖遺傳
* 連鎖:若干非等位基因位於同一染色體而發生連鎖遺傳的現象。
*不完全連鎖(部分連鎖):可產生多種配子,後代出現新性狀的組合,但新組合較理論數為少。非等位基因完全連鎖的情形少見,一般是不完全連鎖。
連鎖遺傳普遍存在於真核生物和原核生物。例如使玉米子粒糊粉層成為有色的基因C為無色基因 c的顯性,使子粒胚乳飽滿發育的基因Sh為凹陷不飽滿基因sh的顯性。
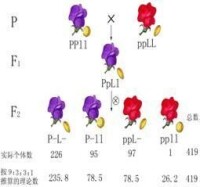
將使有色、飽滿的(CCShSh)與無色、凹陷的(ccshsh)兩個純合親本雜交, (CcShsh)再與無色、凹陷(ccshsh)的親本回交(測交),產生8368粒測交子粒(它們雖然長在F1果穗上,但實際上已是測交子代Ft),
其中有4032粒是有色、飽滿的,4035粒是無色、凹陷的,兩者分別與雙親的子粒性狀相似;另有149粒是有色、凹陷的,152粒是無色、飽滿的,兩者都屬於雙親子粒性狀的重組型。所以F1測交產生的親型測交子代為,重組型測交子代為。出現以上雜交和測交結果的機制是:
①測交後代中親型出現的頻率多達96.4%,說明親本之一的 C與Sh在從親代對測交子代的傳遞過程中有伴隨不分的趨勢,親本之二的 c和sh也表現同樣的趨勢。造成這種趨勢的原因在於C-c和Sh-sh兩對基因是在1對同源染色體的不同位點上;前一親本的基因型為CCShSh,后一親本的基因型為ccshsh,因而F1的基因型為CcShsh。照理, 要產生CSh和csh兩種親型配子,它們與測交親本的csh配子受精結合,會出現和兩種與雙親表型一樣的測交子代。
②測交後代中出現3.6%的重組型 個體,是因為 的少數孢母細胞在產生配子的減數分裂過程中,這對染色體在C-c和Sh-sh之間發生了非姊妹染色單體的交叉互換,從而產生Csh和cSH兩種重組型配子,它們與測交親本的csh配子受精結合,產生和兩種重組型測交子代。
交換髮生的過程是:如圖所示, 的某一孢母細胞內,C-c與Sh-sh之間,如果發生非姊妹染色單體的交換,可產生50%重組型配子和50%親型配子。既然上述測交子代中的兩種重組型子粒只佔3.6%,說明F1所產生的兩種重組型配子(Csh和cSH)也是3.6%,換言之,F1有2×3.6%的孢母細胞,交換正好發生在 C-c與Sh-sh之間。
在使F1與隱性純合體測交的情況下, 產生配子時的交換率(交換 值)與測交子代表型的重組率(重組值)是完全一致的。
當的全部孢母細胞的交換都發生在某2對連鎖基因的連鎖區之內時,重組率為50%;當F1的全部孢母細胞的交換都發生在某2對連鎖基因的連鎖區之外時,重組率為0。但這兩種極端的情形都很少發生,所以重組率總是大於0和小於 50%。
連鎖基因在染色體上的位點相距越遠,則在它們之間發生交換的機率越大,重組率就趨向高值;反之就趨向低值。所以遺傳學中就以某2個連鎖基因之間重組值的大小,表示它們之間相對距離的遠近,並據此繪製連鎖遺傳圖譜。如C-c和Sh-sh是在玉米第9染色體的短臂上,決定玉米子實為非糯質的基因(WX)和糯質的基因(wX)與C-c和Sh-sh也是連鎖的,其重組率分別為22.3%和18.7%
連鎖遺傳
1900年孟德爾遺傳規律被重新發現后,人們以更多的動植物為材料進行雜交試驗,其中屬於兩對性狀遺傳的結果,有的符合獨立分配定律,有的不符。摩爾根以果蠅為試驗材料進行研究,最後確認所謂不符合獨立遺傳規律的一些例證,實際上不屬獨立遺傳,而屬另一類遺傳,即連鎖遺傳。於是繼孟德爾的兩條遺傳規律之後,連鎖遺傳成為遺傳學中的第三個遺傳規律。所謂連鎖遺傳定律,就是原來為同一親本所具有的兩個性狀,在中常常有連繫在一起遺傳的傾向,這種現象稱為連鎖遺傳。
連鎖遺傳定律的發現,證實了染色體是控制性狀遺傳基因的載體。通過交換的測定進一步證明了基因在染色體上具有一定的距離的順序,呈直線排列。這為遺傳學的發展奠定了堅實地科學基礎。

基本信息
- 中文名
- 連鎖遺傳
- 連鎖
- 若干非等位基因位於同一染色體而發生連鎖遺傳的現象
- 完全連鎖
- 位於同源染色體上非等位基因之間不能發生非姐妹染色單體之間的交換,F1隻產生兩種親型配子,其自交或測交後代個體的表現型均為親本組合
- 性質
- 原來親本所具有的兩個或多個性狀,在F2常有 聯繫在一起遺傳的傾向