β氧化
β氧化
β氧化是指代謝氧化的一個長鏈脂肪酸通過連續周期的反應,再每一步的脂肪酸縮短形成含兩個原子碎片移除乙醯輔酶a的過程。脂肪酸β氧化過程可概括為活化、轉移、β氧化及最後經三羧酸循環被徹底氧化生成CO2和H₂O並釋放能量等。
定義:脂肪酸在一系列酶的作用下,在α碳原子和β碳原子之間斷裂,β碳原子被氧化成羧基,生成含有兩個碳原子的乙醯輔酶A,和較原來少兩個碳原子的脂肪酸。
在肝臟內脂肪酸經β-氧化作用生成乙醯輔酶A,兩分子的乙醯輔酶A可縮合生成乙醯乙酸。乙醯乙酸可脫羧生成丙酮,也可還原生成β-羥丁酸。乙醯乙酸、β-羥丁酸和丙酮總稱為酮體。肝臟不能利用酮體,必須經血液運至肝外組織特別是肌肉和腎臟,再轉變為乙醯輔酶A而被氧化利用。酮體作為有機體代謝的中間產物,在正常的情況下,其產量甚微,患糖尿病或食用高脂肪膳食時,血中酮體含量增高.尿中也能出現酮體。
脂肪酸是由一條長的烴基上附加一個羧基的化合物,溶解度一般不大,主要來源於脂肪在人體消化道內的水解。
碳原子個數為偶數的脂肪酸進入人體后,其羧基在細胞質基質中與乙醯輔酶A(乙醯CoA)結合,之後循環往複地被催化脫去乙基,產生新的乙醯CoA,直至碳原子全部脫去。
β氧化
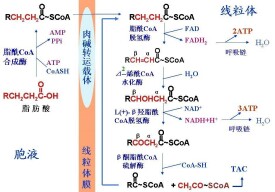
脂肪酸的活化在胞液中進行,而催化脂肪酸氧化分解的酶系存在於線粒體的基質內,因此活化的脂醯CoA必須進入線粒體內才能代謝。長鏈的脂醯CoA不能直接透過線粒體內膜,需依靠特殊的運送機制將它們轉運進入線粒體。肉(毒)鹼[carnitine,L-(CH3)3N+CH2CH(OH)CH2COO-,L-β羥-γ-三甲氨基丁酸]是脂醯基的轉運載體。
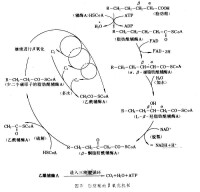
脂醯CoA在線粒體基質中,被疏鬆結合在一起的脂酸β-氧化多酶複合體催化,脂醯基的β碳原子發生氧化,經脫氫、水化、再脫氫、硫解4步連續反應,生成1分子乙醯CoA和1分子比原來少2個碳原子的脂醯CoA。
1.脂肪酸的活化——脂醯CoA的生成
脂肪酸的活化反應在胞液中進行,脂肪酸在脂醯CoA合成酶(acyl-CoA synthetase)催化下,在ATP、CoA-SH、Mg2+ 存在的條件下,活化為脂醯CoA。脂醯CoA含有高能硫酯鍵,而且水溶性增大,使脂醯基的代謝活性明顯增加。分子中的CoA是脂醯基的載體。由於反應過程中生成的焦磷酸(PPi),迅速被細胞內的焦磷酸酶水解,阻止了逆向反應的發生,因此1分子脂肪酸活化成脂醯CoA,實際上消耗了2個高能磷酸鍵。
2.脂醯CoA進入線粒體
脂肪酸的活化在胞液中進行,而催化脂肪酸氧化分解的酶系存在於線粒體基質,因此活化的脂醯CoA必須進入線粒體才能分解。脂醯CoA不能直接透過線粒體內膜,其脂醯基需經肉(毒)鹼轉運才能進入基質。線粒體內膜的兩側存在著肉毒鹼脂醯轉移酶Ⅰ及Ⅱ,在位於線粒體內膜外側面的酶Ⅰ的催化下,脂醯CoA轉化為脂肪醯肉毒鹼,而移到膜內側,進入膜內側的脂肪醯肉毒鹼又經酶Ⅱ的催化而重新轉變成脂醯CoA,並釋放出肉毒鹼(圖5-1-8)。肉毒鹼脂醯轉移酶Ⅰ是限速酶,脂醯CoA進入線粒體是脂肪酸氧化的限速步驟,當飢餓、高脂低糖膳食或糖尿病時,體內糖利用發生障礙,需要脂肪酸供能,這時肉毒鹼脂醯轉移酶Ⅰ活性增加,脂肪酸氧化增強。
1. 脫氫 脂醯CoA在脂醯CoA脫氫酶催化下,α、β碳原子各脫下一個氫原子生成反△2烯脂醯CoA。FAD接受這對氫原子生成FADH2。
2. 水化 反△2烯脂醯CoA在△2烯脂醯水化酶的催化下,加水生成L(+)-β-羥脂醯CoA。
3. 再脫氫 L(+)-β-羥脂醯CoA在β-羥脂醯CoA脫氫酶的催化下,在β碳原子上脫去2個氫原子,生成β-酮脂醯CoA,NAD+接受脫下的這對氫原子生成NADH+H+。
4. 硫解 β-酮脂醯CoA在β-酮脂醯CoA硫解酶的催化下,α與β碳原子間發生斷裂,1分子CoASH參與反應,生成1分子乙醯CoA和少了2個碳原子的脂醯CoA。
以上生成的比原來少2個碳原子的脂醯CoA,可再進行β-氧化,如此反覆進行,直至生成4碳的丁醯CoA,後者進行最後一次β-氧化,將1分子脂醯CoA全部分解為乙醯CoA,完成β-氧化全過程。
脂肪酸經β-氧化生成的乙醯CoA,在線粒體內可與其他代謝途徑生成的乙醯CoA一起進入三羧酸循環,充分氧化成水和CO2,還可以轉變為其他代謝中間產物(如酮體、膽固醇等)
1.β-氧化過程在線粒體基質內進行;
2.β-氧化為一循環反應過程,由脂肪酸氧化酶系催化,反應不可逆;
3.需要FAD,NAD,CoA為輔助因子;
4.每循環一次,生成一分子FADH2,一分子NADH,一分子乙醯CoA和一分子減少兩個碳原子的脂醯CoA。
5.脂肪酸氧化分解是體內重要的能量來源。
軟脂醯CoA需經7次β-氧化,分解生成8分子乙醯CoA、7FADH2、7NADH+H+。因此1分子軟脂醯CoA氧化共產生(7×2)+(7×3)+(8×12)=131ATP,減去軟脂酸活化時消耗的2個高能磷酸鍵(相當於2個ATP),凈生成129分子ATP。
β氧化作用的提出是在二十世紀初,Franz Knoop 在此方面作出了關鍵性的貢獻。他將末端甲基上連有苯環的脂肪酸喂飼狗,然後檢測狗尿中的產物。結果發現,食用含偶數碳的脂肪酸的狗的尿中有苯乙酸的衍生物苯乙尿酸,而食用含奇數碳的脂肪酸的狗的尿中有苯甲酸的衍生物馬尿酸。 Knoop由此推測無論脂肪酸鏈的長短,脂肪酸的降解總是每次水解下兩個碳原子。據此,Knoop 提出脂肪酸的氧化發生在β-碳原子上,而後Ca與Cb之間的鍵發生斷裂,從而產生二碳單位,此二碳單位Knoop推測是乙酸。
以後的實驗證明Knoop推測的準確性,由此提出了脂肪酸的β-氧化作用。
β-氧化作用是指脂肪酸在β-碳原子上進行氧化,然後α-碳原子和β-碳原子之間鍵發生斷裂。每進行一次β-氧化作用,分解出一個二碳片段,生成較原來少兩個碳原子的脂肪酸。
後來對CoA的發現以及分離和提純了參與脂肪酸氧化的各種酶,更弄清了其氧化機制的細節。E.P.Kennedy 和 A.L.Lehninger(1949)指出此氧化系統存在於線粒體中,後來D.E.Green及F.Lynen(1953)各自獨立地從線粒體的丙酮粉末提取出可溶性酶,成功地分離出β氧化各個階段的酶,明確了脂肪β氧化,按下述過程進行:
(1)由脂肪酸活化酶使脂肪酸與 CoA結合,
(2)由乙醯CoA脫氫酶的作用使乙醯CoA脫氫,
(3)由烯醯CoA水合酶的作用使烯醯CoA加水,
(4)由β-羥基乙醯 CoA脫氫酶的作用使β-羥基乙醯 CoA脫氫,
(5)由β-酮醯CoA硫解酶的作用使β酮醯CoA裂解。
經以上5個階段逐次遊離出來的乙 醯CoA(C2片段)經三羧酸循環而氧化。其能量收支為每分子棕櫚酸(C16)產生130分子ATP。不飽和脂肪酸的氧化除需上述各種酶之外,還需要催化3-順-烯醯CoA轉變成2-反式的3-順, 2-反-烯醯CoA異構酶和催化D(一)-3-羥式成L(+)-3-羥式的3-羥乙醯CoA-3-表異構酶參與。由奇數C原子脂肪酸分解產生的丙醯CoA,通過羧化及異構化而轉變成琥珀醯CoA再進一步變化。
基本信息
- 中文名
- β氧化
- 外文名
- β-oxidation
- 發現人
- Franz Knoop