三羧酸循環
生物化學術語
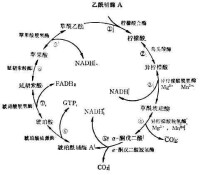
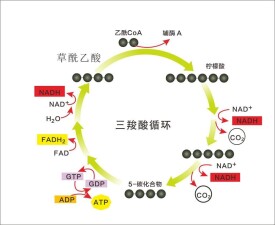
三羧酸循環(tricarboxylic acid cycle,TCA cycle)是需氧生物體內普遍存在的代謝途徑。原核生物中分佈於細胞質,真核生物中分佈在線粒體。因為在這個循環中幾個主要的中間代謝物是含有三個羧基的有機酸,例如檸檬酸(C6),所以叫做三羧酸循環,又稱為檸檬酸循環(citric acid cycle)或者是TCA循環;或者以發現者Hans Adolf Krebs(英1953年獲得諾貝爾生理學或醫學獎)的姓名命名為Krebs循環。三羧酸循環是三大營養素(糖類、脂類、氨基酸)的最終代謝通路,又是糖類、脂類、氨基酸代謝聯繫的樞紐。
檸檬酸循環(citric acid cycle):也稱為三羧酸循環(tricarboxylic acid cycle,TCA循環,TCA),Krebs循環。是用於將乙醯CoA中的乙醯基氧化成二氧化碳和還原當量的酶促反應的循環系統,該循環的第一步是由乙醯CoA與草醯乙酸縮合形成檸檬酸。反應物乙醯輔酶A(Acetyl-CoA)(一分子輔酶A和一個乙醯相連)是糖類、脂類、氨基酸代謝的共同的中間產物,進入循環後會被分解最終生成產物二氧化碳併產生H,H將傳遞給輔酶I--尼克醯胺腺嘌呤二核苷酸(NAD+) (或者叫煙醯胺腺嘌呤二核苷酸)和黃素腺嘌呤二核苷酸(FAD),使之成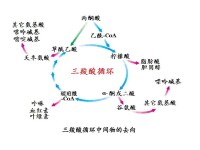 為NADH + H+和FADH2。 NADH + H+ 和 FADH2 攜帶H進入呼吸鏈,呼吸鏈將電子傳遞給O2產生水,同時偶聯氧化磷酸化產生ATP,提供能量。
為NADH + H+和FADH2。 NADH + H+ 和 FADH2 攜帶H進入呼吸鏈,呼吸鏈將電子傳遞給O2產生水,同時偶聯氧化磷酸化產生ATP,提供能量。
三羧酸循環
真核生物的線粒體基質和原核生物的細胞質是三羧酸循環的場所。它是呼吸作用過程中的一步,之後高能電子在NAHD+H+和FADH2的輔助下通過電子傳遞鏈進行氧化磷酸化產生大量能量。
糖類物質如葡萄糖或糖原在有氧條件下徹底氧化,產生二氧化碳和水,並釋放出能量的過程稱為糖的有氧氧化。人們發現,肌肉糜在有氧存在時,沒有乳酸的生成,也沒有丙酮酸的累積,但仍有能量放出,為什麼?著名生物化學家H.Kreb等為闡明在有氧情況下丙酮酸的代謝,作了大量的研究工作,提出了糖的有氧氧化途徑,為此獲1953年諾貝爾獎。
糖的有氧氧化與糖的無氧酵解有一段共同途徑,即葡萄糖一丙酮酸,所不同的是在生成丙酮酸以後的反應。在有氧情況下,丙酮酸在丙酮酸脫氫酶系的催化下,氧化脫羧生成乙醯CoA,後者再經三羧酸循環(tricarboxylic acid cycle)氧化成CO2,和H2O。
在有氧情況下,肌糖原酵解的產物乳酸也可能轉變成丙酮酸。例如,血乳酸可被心肌等組織利用作為能源,是人體在激烈運動后的恢復期所進行的一個反應。在這段恢復時間,呼吸仍加快加深,乳酸重新氧化成丙酮酸,後者再進一步氧化成水和CO2 。
克雷布斯博士在第二次世界大戰爆發期間因受到納粹的迫害,不得不逃往英國。雖然在德國,他是位非常優秀的醫生,但是在英國,由於沒有行醫許可證,得不到社會的承認,他只能轉而從事基礎醫學的研究。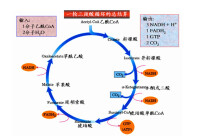
三羧酸循環
剛開始選擇課題時,僅僅因為他對食物在體內究竟是如何變成水和二氧化碳這一課題充滿了興趣,他便毫不猶豫地選擇了這個課題,並且著手調查前人研究這一課題的各種材料。在報告中,他看到有的學者報告說:“A物質經過氧化變成了B物質。”又有學者說:“C物質經過氧化變成了D物質,然後又進一步變成E物質。”還有學者認為:“C物質是從B物質中得到的。或者可以說,是F物質變成了G物質。”另外一些學者則認為,是“G物質經過氧化變成A物質”等等。看著來自四面八方的研究報告,克雷布斯想,如果把這些零散的數據整理出來,說不定可以發現食物代謝的結構。就像玩解謎遊戲那樣,克雷布斯將這些數據仔細整理了一番,結果發現食物在體內是按F、G、A、B、C、D、E這樣一個順序變化的。再仔細了解從A到F這些化學物質,發現E和F之間斷了鏈。如果E和F之間存在一種X物質,那麼,這條食物循環反應鏈就完整了。馬上集中精力,全力尋找X物質。4年後終於查明,X物質就是如今放在飲料中作為酸味添加劑的檸檬酸。他完成了食物的循環鏈,並且將它命名為檸檬酸循環。克雷布斯的循環理論解釋了食物在體內進入檸檬酸循環后,按照A、B、C、D、E、X、F、G的順序循環反應,最終氧化成二氧化碳和水。他的偉大不僅僅在於發現了幾個化學物質的變化,而且在於將每一個活的變化整理出來,找出了可以解釋動態生命現象的結構。由於這一業績,他在1953年獲諾貝爾生理學或醫學獎。檸檬酸循環也叫三羧酸循環或TCA循環。進入體內的營養成分在糖酵解→檸檬酸循環→電子傳遞等一系列呼吸作用下得到分解,產生能量。
三羧酸循環(tricarboxylic acid cycle )是一個由一系列酶促反應構成的循環反應系統,在該反應過程中,首先由乙醯輔酶A(C2)與草醯乙酸(OAA)(C4)縮合生成含有3個羧基的檸檬酸(C6),經過4次脫氫(3分子NADH+H+和1分子FADH2),1次底物水平磷酸化,最終生成2分子CO2,並且重新生成草醯乙酸的循環反應過程。
化學反應
Acetyl-CoA + 3 NAD + FAD + GDP + P + 2 HO →CoA-SH + 3 NADH + 3 H + FADH + GTP + 2 CO
兩個碳原子以CO的形式離開循環。循環最後草醯乙酸會再次生成,再次從乙醯輔酶A中得到兩個碳原子。就是說,一分子六碳化合物(檸檬酸)經過多部反應分解成一分子四碳化合物(草醯乙酸)。草醯乙酸會在接下來的反應中遵循同樣的途徑獲得兩個碳原子,再次成為檸檬酸。
能量會在接下來的其中一步反應里以GTP的形式釋放(和ATP一樣,是細胞的能量貨幣)。但是循環中生成的氫載體(NADH + H and FADH)將會在細胞呼吸鏈里釋放更多的能量,這也正是細胞呼吸的主要目的。檸檬酸循環的前提是,早先進行的糖酵解等過程能提供足夠的活化乙酸,以乙醯輔酶A的形式出現在循環。NADH + H 和 FADH是輔酶,它們能攜帶質子和電子,並在需要的時候釋放它們。
循環中產生的總能量為一分子ATP(準確來說是:GTP),而細胞呼吸的全部四步反應(包括呼吸鏈中的內呼吸),一個葡萄糖分子則產生32分子的ATP。2002年之前一直認為是38ATP,當時認為一個FADH2可以產生2個ATP,一個NADH2可以產生3個ATP,這是理想化化學計算的結果。實測一個FADH2可以產生1.5個ATP,一個NADH2可以產生2.5個ATP。詳情請查閱電子傳遞鏈與氧化磷酸化。
如進行蘋果酸穿梭則不會減少能量,還是32ATP,在腦等部位會進行3磷酸甘油穿梭,減少2分子ATP,最終凈產生30ATP。所以說,在生物化學專業答題時需回答32或30。
![三羧酸循環[生物化學術語]](https://i1.twwiki.net/cover/w200/m2/8/m28868f726e8f71fb33b410c3ff5c1dcc.jpg)
三羧酸循環[生物化學術語]
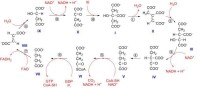
1、乙醯-CoA進入三羧酸循環
乙醯CoA具有硫酯鍵,乙醯基有足夠能量與草醯乙酸的羧基進行醛醇型縮合。首先檸檬酸合酶的組氨酸殘基作為鹼基與乙醯-CoA作用,使乙醯-CoA的甲基上失去一個H+,生成的碳陰離子對草醯乙酸的羰基碳進行親核攻擊,生成檸檬醯-CoA中間體,然後高能硫酯鍵水解放出遊離的檸檬酸,使反應不可逆地向右進行。該反應由檸檬酸合酶(citratesynthase)催化,是很強的放能反應。由草醯乙酸和乙醯-CoA合成檸檬酸是三羧酸循環的重要調節點,檸檬酸合酶是一個變構酶,ATP是檸檬酸合酶的變構抑製劑,此外,α-酮戊二酸、NADH能變構抑制其活性,長鏈脂醯-CoA也可抑制它的活性,AMP可對抗ATP的抑制而起激活作用。
2、異檸檬酸形成
![三羧酸循環[生物化學術語]](https://i1.twwiki.net/cover/w200/m1/d/m1db5df7aeaa4321b64844c821683d402.jpg)
三羧酸循環[生物化學術語]
3、第一次脫氫——異檸檬酸脫氫酶
在異檸檬酸脫氫酶作用下,異檸檬酸的仲醇氧化成羰基,生成草醯琥珀酸(oxalosuccinicacid)的中間產物,後者在同一酶表面,快速脫羧生成α-酮戊二酸(α-ketoglutarate)、NADH和CO2,此反應為β-氧化脫羧,此酶需要鎂離子作為激活劑。此反應是不可逆的,是三羧酸循環中的限速步驟,ADP是異檸檬酸脫氫酶的激活劑,而ATP,NADH是此酶的抑製劑。
4、第二次脫氫——α-酮戊二酸脫氫酶
在α-酮戊二酸脫氫酶系作用下,α-酮戊二酸氧化脫羧生成琥珀醯-CoA、NADH·H+和CO₂,反應過程完全類似於丙酮酸脫氫酶系催化的氧化脫羧,屬於α-氧化脫羧,氧化產生的能量中一部分儲存於琥珀醯coa的高能硫酯鍵中。α-酮戊二酸脫氫酶系也由三個酶(α-酮戊二酸脫羧酶、硫辛酸琥珀醯基轉移酶、二氫硫辛酸脫氫酶)和五個輔酶(tpp、硫辛酸、hscoa、NAD+、FAD)組成。此反應也是不可逆的。α-酮戊二酸脫氫酶複合體受ATP、GTP、NADH和琥珀醯-CoA抑制,但其不受磷酸化/去磷酸化的調控。
5、底物磷酸化生成ATP
![三羧酸循環[生物化學術語]](https://i1.twwiki.net/cover/w200/mb/7/mb78289b1d8bd19c86f3485603427f67c.jpg)
三羧酸循環[生物化學術語]
6、第三次脫氫——琥珀酸脫氫酶
琥珀酸脫氫酶(succinatedehydrogenase)催化琥珀酸氧化成為延胡索酸。該酶結合在線粒體內膜上,而其他三羧酸循環的酶則都是存在線粒體基質中的,這酶含有鐵硫中心和共價結合的FAD,來自琥珀酸的電子通過FAD和鐵硫中心,然後進入電子傳遞鏈到O₂,丙二酸是琥珀酸的類似物,是琥珀酸脫氫酶強有力的競爭性抑制物,所以可以阻斷三羧酸循環。
7、延胡索酸的水化
延胡索酸酶僅對延胡索酸的反式雙鍵起作用,而對順丁烯二酸(馬來酸)則無催化作用,因而是高度立體特異性的。
8、第四次脫氫——蘋果酸脫氫酶(草醯乙酸再生)
在蘋果酸脫氫酶(malicdehydrogenase)作用下,蘋果酸仲醇基脫氫氧化成羰基,生成草醯乙酸(oxalocetate),NAD+是脫氫酶的輔酶,接受氫成為NADH·H+(圖4-5)。
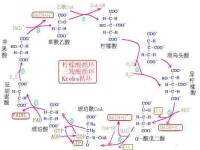
三羧酸循環
乙醯-CoA+3NAD++FAD+ADP+Pi+CoA-SH—→2CO2+3NADH+FADH2+ATP+3H++CoA-SH
1、CO₂的生成,循環中有兩次脫羧基反應(反應3和反應4)兩次都同時有脫氫作用,但作用的機理不同,由異檸檬酸脫氫酶所催化的β氧化脫羧,輔酶是nad+,它們先使底物脫氫生成草醯琥珀酸,然後在Mn2+或Mg2+的協同下,脫去羧基,生成α-酮戊二酸。α-酮戊二酸脫氫酶系所催化的α氧化脫羧反應和前述丙酮酸脫氫酶系所催經的反應基本相同。應當指出,通過脫羧作用生成CO₂,是機體內產生CO₂的普遍規律,由此可見,機體CO₂的生成與體外燃燒生成Co2的過程截然不同。
![三羧酸循環[生物化學術語]](https://i1.twwiki.net/cover/w200/me/4/me4f0d10897ebf6a854045e095a6e617b.jpg)
三羧酸循環[生物化學術語]
3、乙醯-CoA中乙醯基的碳原子,乙醯-CoA進入循環,與四碳受體分子草醯乙酸縮合,生成六碳的檸檬酸,在三羧酸循環中有二次脫羧生成2分子CO₂,與進入循環的二碳乙醯基的碳原子數相等,此時乙醯輔酶A中的2個碳已全部轉變為CO₂,同時其中的一部分能量已轉變成了NADH和ATP中的能量。
4、三羧酸循環的中間產物,從理論上講,可以循環不消耗,但是由於循環中的某些組成成分還可參與合成其他物質,而其他物質也可不斷通過多種途徑而生成中間產物,所以說三羧酸循環組成成分處於不斷更新之中。
下面以轉氨基作用偶聯尿素循環為例,TCA的中間產物可以作為其他代謝途徑的前體。
例如草醯乙酸——→天冬氨酸(Asp)
α-酮戊二酸——→谷氨酸(Glu)
草醯乙酸——→丙酮酸——→丙氨酸(Ala)
其中丙酮酸羧化酶催化的生成草醯乙酸的反應最為重要。因為草醯乙酸的含量多少,直接影響循環的速度,因此不斷補充草醯乙酸是使三羧酸循環得以順利進行的關鍵。三羧酸循環中生成的蘋果酸和草醯乙酸也可以脫羧生成丙酮酸,再參與合成許多其他物質或進一步氧化。
三羧酸循環
丙酮酸脫氫酶複合體受別構調控也受化學修飾調控,該酶複合體受它的催化產物ATP、乙醯-CoA和NADH有力的抑制,這種別構抑制可被長鏈脂肪酸所增強,當進入三羧酸循環的乙醯-CoA減少,而AMP、CoA和NAD+堆積,酶複合體就被別構激活,除上述別位調節,在脊椎動物還有第二層次的調節,即酶蛋白的化學修飾,PDH含有兩個亞基,其中一個亞基上特定的一個絲氨酸殘基經磷酸化后,酶活性就受抑制,脫磷酸化活性就恢復,磷酸化-脫磷酸化作用是由特異的磷酸激酶和磷酸蛋白磷酸酶分別催化的,它們實際上也是丙酮酸酶複合體的組成,即前已述及的調節蛋白,激酶受ATP別構激活,當ATP高時,PDH就磷酸化而被激活,當ATP濃度下降,激酶活性也降低,而磷酸酶除去PDH上磷酸,PDH又被激活了。
對三羧酸循環中檸檬酸合成酶、異檸檬酸脫氫酶和α-酮戊二酸脫氫酶的調節,主要通過產物的反饋抑制來實現的,而三羧酸循環是機體產能的主要方式。因此ATP/ADP與NADH/NAD+兩者的比值是其主要調節物。ATP/ADP比值升高,抑制檸檬酸合成酶和異檸檬酶脫氫酶活性,反之ATP/ADP比值下降可激活上述兩個酶。NADH/NAD+比值升高抑制檸檬酸合成酶和α-酮戊二酸脫氫酶活性,除上述ATP/ADP與NADH/NAD+之外其它一些代謝產物對酶的活性也有影響,如檸檬酸抑制檸檬酸合成酶活性,而琥珀醯-CoA抑制α-酮戊二酸脫氫酶活性。總之,組織中代謝產物決定循環反應的速度,以便調節機體ATP和NADH濃度,保證機體能量供給。
TCA的生物學意義可以分為兩方面論述,1.能量代謝 2.物質代謝
1.三羧酸循環是機體將糖或其他物質氧化而獲得能量的最有效方式。在糖代謝中,糖經此途徑氧化產生的能量最多。毎分子葡萄糖經有氧氧化生成H2O和CO2時,可凈產生32分子ATP或30分子ATP。
2.三羧酸循環是糖、脂,蛋白質,甚至核酸代謝,聯絡與轉化的樞紐。
(1)此循環的中間產物(如草醯乙酸、α-酮戊二酸)是合成糖、氨基酸、脂肪等的原料。
其中OAA可以脫羧成為PEP,參與糖異生,重新合成生物體內的能源。acetylCOA可以合成丙二醯ACP,參與軟脂酸合成。OAA可以在轉氨酶的參與下,進行轉氨基作用,生成Asp,參與urea cycl,合成精氨酸代琥珀酸等尿素前體物質。其中某些代謝物質,還能參與嘌呤和嘧啶的合成,甚至合成卟啉ring,參與血紅蛋白合成。
(2)TCA是糖、蛋白質和、和脂肪徹底氧化分解的共同途徑:蛋白質的水解產物(如谷氨酸、天冬氨酸、丙氨酸等脫氨后或轉氨后的碳架)要通過三羧酸循環才能被徹底氧化,產生大量能量;脂肪分解后的產物脂肪酸經β-氧化後生成乙醯CoA以及甘油,甘油經過EMP途徑也生成乙醯CoA,最終也要經過三羧酸循環而被徹底氧化。糖代謝的所有途徑最後生成Pyruvate,脫氫成為acetyl-CoA,參與TCA。
綜上所述,三羧酸循環是聯繫三大物質代謝的,也是能量代謝的樞紐。
1、為機體提供能量:每摩爾葡萄糖徹底氧化成H2O和CO2時,凈生成30mol或32mol(糖原則生成31~ 33mol)ATP。因此在一般生理條件下,各種組織細胞(除紅細胞外)皆從糖的有氧氧化獲得能量。糖的有氧氧化不但產能效率高,而且逐步釋能,並逐步儲存於ATP分子中,因此能的利用率也極高。
2、三羧酸循環是三大營養物質的共同氧化途徑:乙醯CoA,不但是糖氧化分解的產物,也是脂肪酸和氨基酸代謝的產物,因此三羧酸循環實際上是三大有機物質在體內氧化供能的共同主要途徑。據估計人體內2/3的有機物質通過三羧酸循環而分解。
3、三羧酸循環是三大物質代謝聯繫的樞紐:糖有氧氧化過程中產生的α-酮戊二酸、丙酮酸和草醯乙酸等與氨結合可轉變成相應的氨基酸;而這些氨基酸脫去氨基又可轉變成相應的酮酸而進入糖的有氧氧化途徑。同時脂類物質分解代謝產生的甘油、脂肪酸代謝產生的乙醯CoA也可進入糖的有氧氧化途徑進行代謝。
基本信息
- 中文名
- 三羧酸循環
- 外文名
- The tricarboxylic acid cycle
- 別名
- 檸檬酸循環
- 類型
- 是糖類、脂類、氨基酸代謝聯繫的樞紐
- 領域
- 生物化學
- 簡稱
- TCA循環