圓口綱
圓口綱
圓口綱(Cyclostomata),魚形動物中最早的一類,無成對偶肢和上下頜的低等脊椎動物,大約起源於4.8億年前。
身體裸露無鱗,呈鰻形。全為軟骨。無偶鰭。無肩帶和腰帶。無上、下頜,所以又稱無頜類(Agnatha)。具1鼻孔。鰓呈囊狀,又稱囊鰓類。舌肌發達,上附角質齒。舌以活塞式運動舐刮魚肉。脊索終生存在。內耳半規管1~2個。生活于海洋或淡水中,有些種類具有洄遊(migration)。
在古生代奧陶紀和志留紀、泥盆紀地層中發現的甲胄魚類化石和現存圓口綱有許多共同之處,如無上、下頜,無偶鰭,有鰓籠和單個鼻孔,內耳有2個半規管等,圓口綱可能是甲胄魚類的後裔,也有人認為它們來自共同的無頜類祖先。
現存的圓口綱動物約70多種,分屬於2個目:七鰓鰻目(Petromyzoniformes)和盲鰻目(Myxiniformes)。代表種如日本七鰓鰻(Lampetra japonicus)、盲鰻(Myxine glutinosa)。營寄生或半寄生生活,以大型魚類及海龜類為寄主。七鰓鰻主要用前端的口漏斗吸附於寄主體表,用角質齒銼破皮膚吸血食肉;盲鰻更能由魚鰓部鑽入寄主體內,吃盡其內臟,使之僅存軀殼,因而常給漁業造成危害。
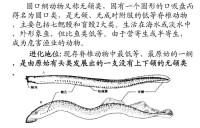
圓口綱
皮膚:皮膚裸露,富含單細胞腺體,能分泌黏液潤滑體表,無鱗片,身體兩側各有1行小孔,是水棲動物的側線器官。
骨骼:骨骼結構原始,由軟骨和結締組織組成,沒有硬骨。脊索成為支持身體的中軸,脊索的背方神經管兩旁有許多小軟骨片,代表了雛形脊椎骨。頭骨構造也很原始,腦顱為一個不完全的軟骨顱,腦頂部僅覆蓋結締組織膜。支持鰓囊的鰓籠由9對橫行的軟骨弧和4對縱行的軟骨條相連接而成。
肌肉:肌肉很原始,體壁肌肉有“W”形的分節現象。
消化系統:口位於口吸盤的底部,由口通入口腔,口腔後為咽,咽分化出2條管道,背方的為食道,腹方的為呼吸道。七鰓鰻無胃分化,食道直接和腸相連,腸內有縱向的螺旋瓣,以增加消化和吸收的面積,腸末端為肛門。消化腺有發達的肝臟,胰臟不發達。
呼吸系統:咽分出腹面的盲管為呼吸管,其兩側各有7個內鰓孔,每個內鰓孔各與一鰓囊相通,鰓囊內有鰓絲,上有毛細血管,能進行氣體交換。
循環系統:基本上和文昌魚相似,但已出現心臟,心臟具有一心房、一心室和一個靜脈竇。
排泄系統:有狹長的腎臟1對,由腹膜固著於體腔壁上,還有2條輸尿管沿體腔後行,開口於泄殖竇。七鰓鰻成體的腎臟從發生上屬中腎,但幼體同時存在前腎和中腎。
神經系統:還很原始。腦雖然分化為端腦、間腦、中腦、小腦和延腦5部分,但體積很小。腦神經10對。
感覺器官、嗅覺器官:有鼻孔,內通往嗅囊,其上有嗅覺細胞。視覺器官為1對眼。聽覺器官有1對內耳,只有前、后2個半規管。味覺器官是口腔的味蕾。
生殖系統:七鰓鰻為雌雄異體,只有單個的精巢或卵巢,但無輸出管道,成熟的精子或卵子落到腹腔。
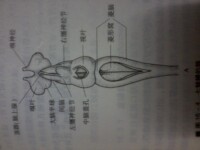
七鰓鰻的腦結構

圓口綱

圓口綱
七鰓鰻目( Petromyzoniformes):約26種。分佈於淡水或海洋,成體營半寄生生活。有口漏斗和角質齒,鰓囊7對,內耳有兩個半規管,雌雄異體發育有變態。如海七鰓鰻( Petromyzon marinus)產於大西洋中,有溯河產卵習性。而東北七鰓鰻(Lαπ petra mori)、日本七鰓鰻(.jφoπicus)、雷氏七鰓鰻(L. reissηeri)等分佈於我國東北松花江、黑龍江等水域,日本七鰓鰻成體在海中生活。東北七鰓鰻兩背鰭分開,板齒9~1枚;本七鰓鰻兩背鰭分開,板齒δ~7枚;雷氏七鰓鰻兩背鰭相連。
盲鰻目( Morini formes):約21種。全為海產,幼體以多毛類為食,也襲擊魚類,鑽入魚體或其他動物體內,吸食其血肉和內臟,營全寄生生活。無口漏斗,口緣有4對觸鬚。鰓囊6~15對。但有些屬的出鰓水管癒合成一個管,在每側有一向外開口的孔內耳1個半規管(實際上,內耳兩個半規管互相連接,看起來如同一個)。雌雄同體,發育無變態。如大西洋盲鰻( Myxine glutinosa),蒲氏黏盲鰻(Eφ tatretus burgeri),紫黏盲鯪。
圓口綱動物有一定的經濟價值,七鰓鰻和盲鰻類均可食用,七鰓鰻有一定的捕撈價值。盲鰻的數量較多,營寄生生活,一般在晚上襲擊魚類,多從鰓部鑽入體腔攝食寄主的內臟和肌肉。對魚類危害較大,但它們也食腐肉,有一定的凈化水質的作用。

基本信息
- 中文名
- 圓口綱
- 界
- 動物界
- 門
- 脊椎動物門
- 亞門
- 脊椎動物亞門
- 綱
- 圓口綱
- 目
- 七鰓鰻目和盲鰻目
- 體征
- 身體裸露無鱗,呈鰻形
- 亞綱
- 七鰓鰻亞綱、盲鰻亞綱