水龍獸屬
獸孔目二齒獸下目的屬
水龍獸屬(學名:Lystrosaurus)意為“鏟子蜥蜴”,是獸孔目二齒獸下目的一屬,存活在二疊紀晚期到三疊紀早期,約2億5000萬年前,化石分佈於南極洲、印度、南非。在1930年代到1970年代曾有許多種被歸類於水龍獸屬,但目前僅有4到6個已承認種。
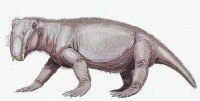
水龍獸是種體型笨重、中等大小的草食性動物,有短胖的四肢,體型接近豬。水龍獸的嘴裡只有兩顆長牙,自上頜延伸出來。上下頜前端可能有喙狀嘴,用來切碎植物。根據肩膀與臀部的關節結構,顯示水龍獸採取往兩側半延展的步態。由於前肢比後肢粗壯,水龍獸被認為有挖掘洞穴的習性。
水龍獸是已知三疊紀早期的最常見陸地脊椎動物,某些該時期的挖掘地點,至少有95%的化石是由水龍獸構成。水龍獸在二疊紀-三疊紀滅絕事件中存活下來的原因,可能是因為它們比其他動物更快適應滅絕事件后的環境;在三疊紀早期,大氣層的氧氣含量低,二氧化碳含量高。近年的研究顯示,水龍獸雖然存活過二疊紀-三疊紀滅絕事件,但三疊紀早期的水龍獸數量,仍然比二疊紀晚期的數量少。
水龍獸是種常見的合弓動物,屬於獸孔目二齒獸下目,體型接近豬,身長約0.9米(3呎),重量約90公斤(200磅)。與其他獸孔目不同,二齒獸類的口鼻部相當短,只有兩顆上頜的長牙。口鼻部的前端可能有角質喙狀嘴,用來切碎植物。水龍獸已具有次生顎。頜部關節原始,只能做出往前、往後的動作,而不能做出往上、往下的動作。水龍獸被推測頜部肌肉佔據頭顱骨頂端、后側的許多空間。眼睛的位置高,角度往前。另外,水龍獸的前肢比後肢粗壯,因此被認為有挖掘洞穴的習性。
有數個特徵顯示水龍獸是採取往兩側半延展的步態:
1、肩胛骨後下端有明顯地骨化,顯示前肢的前後步伐大,也降低身體的側向動作靈活度。
2、薦椎有5節,體積大,但彼此之間、與骨盆之間並未癒合。這使水龍獸的脊椎活動較不易彎曲,當它們行進時身體不會左右擺動。薦椎少於5節的獸孔目,被認為採取四肢往兩側延展的步態,類似現代蜥蜴。恐龍與哺乳類的四肢直立於身體下方,而薦椎彼此之間癒合,薦椎與骨頭之間癒合。
水龍獸
水龍獸的第一個化石,是在19世紀後期由傳教士兼業餘化石挖掘者Elias Root Beadle所發現。Beadle將這化石的消息通知古動物學家奧塞內爾·查利斯·馬什(Othniel Charles Marsh),但馬什沒有回應。馬什的敵手愛德華·德林克·科普(Edward Drinker Cope)獲得這些化石,並在1870年命名為水龍獸。水龍的屬名在古希臘文意為“鏟子蜥蜴”。在1871年,馬什也購得一個頭顱骨,但並沒有詳細地研究。
大陸漂移學說
在1969到1970年,埃德溫·爾伯特(Edwin H. Colbert)與他的團隊在南極縱貫山脈的Coalsack Bluff發現水龍獸的化石,有助於確認大陸漂移學說而說服了懷疑論者。水龍獸目前已在非洲南部、印度、中國的早侏羅紀地層發現。
水龍獸的化石發現於二疊紀晚期到三疊紀早期的陸相地層,大部分來自於非洲,其他化石則發現於印度、中國、蒙古、俄羅斯的歐洲部分、以及南極洲。由於水龍獸的分佈廣泛,因此被當成大陸漂移學說的重要證據之一。
大多數的水龍獸化石發現於南非,尤其集中於卡魯盆地。該地區已發現大量水龍獸化石,也命名了許多種,但科學家對於該地區有幾種的水龍獸,目前還沒有定論。
在1930年代到1970年代之間,水龍獸屬曾經包含23個種。在1980年代到1990年代,則剩下6個已承認種:L. curvatus、L. platyceps、L. oviceps、L. maccaigi、L. murrayi、以及L. declivis。在2006年的新研究,則縮減到4個種,原本的L. platyceps、L. oviceps被併入L. curvatus。
模式種L. murrayi與L. declivis的化石,僅發現於二疊紀晚期的地層。L. maccaigi是體型最大、最特化的一種,而L. curvatus是最原始的一種。同樣發現於南非的Kwazulusaurus,化石類似水龍獸,尤其是L. curvatus。Kwazulusaurus有可能是L. curvatus的祖先,或是其祖先的近親。
1.L. maccaigi的化石僅發現於二疊紀晚期地層,可能沒有存活過二疊紀-三疊紀滅絕事件。由於L. maccaigi是較為衍化的物種,但在化石紀錄的時間很短,沒有可追溯的直系物種,有研究推測它們原本存活於其他二疊紀地層,然後遷徙至卡魯地區。L. curvatus的化石則集中在狹窄的帶狀地區,生存於滅絕事件前後的短暫時期,因此可以當成判斷兩個時代交界的化石之一。在尚比亞的二疊紀晚期地層,曾經發現一個L. curvatus的頭顱骨。由於過去在卡魯盆地的二疊紀地層,沒有發現L. curvatus的地層,曾有理論推測L. curvatus從尚比亞遷徙至南非卡魯盆地。但近年在卡魯盆地的二疊紀地層發現L. curvatus的化石,推翻上述理論。
2.L. georgi的化石發現於俄羅斯的莫斯科地區,地質年代為三疊紀最早期,它們可能是L. curvatus的近親。
在三疊紀最早期的數百萬年,水龍獸是最常見的陸地脊椎動物。在某些該時期的挖掘地點,至少有95%的化石是由水龍獸構成。至少有一個種存活過二疊紀-三疊紀滅絕事件。由於三疊紀早期缺少肉食性動物的獵食,以及其他草食性動物的競爭,水龍獸迅速演化輻射出多個物種,而成為三疊紀早期的最常見陸地脊椎動物。這是地球歷史中的唯一時期,陸地生態系統幾乎由某種動物所佔據。有少數二疊紀物種也存活過二疊紀-三疊紀滅絕事件,例如獸頭亞目的四犬齒獸(Tetracynodon)、麝喙獸(Moschorhinus)、似鼬鱷獸(Ictidosuchoides),但數量沒有水龍獸繁盛。一位研究人員估計陸地脊椎動物花了3000萬年,直到三疊紀晚期,才恢復之前的繁盛與多樣性。
目前有數個理論,試圖解釋水龍獸存活過二疊紀-三疊紀滅絕事件的原因,以及水龍獸迅速成為三疊紀早期優勢物種的原因:
• 二疊紀-三疊紀滅絕事件的原因之一,是大氣層的氧含量下降,二氧化碳含量上升。水龍獸被推測可能是穴居動物,可能因此適應較差的空氣品質,得以存活過這次滅絕事件。水龍獸的多項特徵,顯示它們有良好的呼吸效能,例如桶狀胸腔可以容納較大肺臟,較短的鼻孔通道使呼吸較快,高神經棘使胸部肌肉在擴張、收縮時產生更大應力。但是,與其他二齒獸類的身體比例相比,水龍獸的胸腔並沒有特別大。三疊紀二齒獸類的神經棘更長,神經棘的長度似乎與步態、移動速度、體型較有關係,與呼吸效能較無關係。L. murrayi、L. declivis的數量也比三疊紀早期的穴居動物更多,例如:前棱蜥、三尖叉齒獸。
• 另一理論推測水龍獸是半水生動物,因此受到滅絕事件的波及較小。但是卡魯盆地的三疊紀早期地層,雖然有許多兩棲類化石,但數量仍少於L. murrayi、L. declivis。
• 體型較大的陸地動物,容易因為滅絕事件而滅絕。此理論可以解釋體型較小、較原始的L. curvatus存活過滅絕事件,而體型較大、較衍化的L. maccaigi消失了。
• 水龍獸的存亡,似乎也與所食的植物有關係。二疊紀晚期的叉蕨(Dicroidium,又譯二舌羊齒),在三疊紀早期繁盛;而較大型的舌羊齒(Glossopteris)則消失了。
• 在三疊紀早期的大型掠食動物,例如獸頭亞目的Moschorhinus、主龍形類的原鱷,在化石紀錄的存在時間很短,可能造成水龍獸的繁盛。
水龍獸曾經出現在BBC的電視節目《與巨獸共舞》(Walking with Monsters),節目單位宣稱二疊紀的小型雙齒獸演化成三疊紀的大型水龍獸。但實際上,兩者可能曾經存活於相同時期。此外,水龍獸還出現在電視節目《動物末日》(Animal Armageddon),節目單位試圖解釋水龍獸具有多物種的原因。

基本信息
- 拉丁學名
- Lystrosaurus
- 界
- 動物界
- 亞門
- 脊椎動物亞門
- 門
- 脊索動物門
- 綱
- 合弓綱
- 亞目
- 異齒亞目
- 科
- 水龍獸科
- 屬
- 水龍獸屬