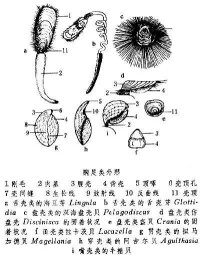
觸手冠,是擬軟體動物包圍口的觸手環的基座。苔蘚蟲類之觸手冠呈簡單的環形,菷蟲類的觸手冠是以口為中心在左右形成複雜的螺旋。腕足綱中的有關節類(Coptotbyris grayi 等)具支持觸手冠的石灰質腕骨,

放大后的觸手冠

觸手冠動物
是擬軟體動物包圍口的觸手環的基座。苔蘚蟲類之觸手冠呈簡單的環形,菷蟲類的觸手冠是以口為中心在左右形成複雜的螺旋。腕足綱中的有關節類(Coptotbyris grayi 等)具支持觸手冠的石灰質腕骨,
觸手冠
附著於背殼。菷蟲類的觸手冠上具有稱為總擔器(lophophoral organ)的功能 不明的腺狀構造。
觸手冠動物(lophophorates)
帚蟲動物、外肛動物及
腕足動物這三類動物系統地位特殊,既有原口動物的特徵又有
后口動物的特徵。因為它們都有一個體壁延伸形成的馬蹄形或環形的觸手冠(lophophore)作為它們的捕食及呼吸器官;而被統稱為“觸手冠動物(lophophorates)”

觸手冠
·均為固著生活,頭部不明顯,神經感官不發達,有發達的外骨骼(介殼、棲管或蟲室)
·都有真體腔,有發達的后腎,併兼作生殖導管,發育中都有一個自由游泳的相似於擔輪幼蟲的幼蟲期。所以,經典的分類把它們列為原口動物一支。
·這類動物也有一些后口動物的典型特徵:

漂亮的觸手冠
觸手冠動物也表現出類似棘皮動物、半索動物等后口動物的三分體區及體腔的體制結構。
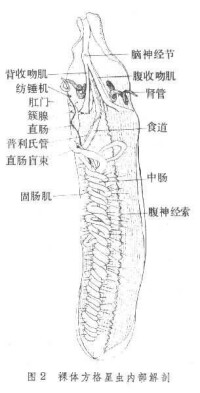
觸手冠
觸手冠動物的
胚胎髮育也表現出某些后口動物的特徵。例如,外肛動物及腕足動物的
卵裂是輻射卵裂而不再是 螺旋卵裂,腕足動物的體腔形成為腸腔法。

觸手冠
在最近50年內,部分或者全部觸手冠動物是后口動物起源的假說得到了越來越多的支持。因此,如何解釋腕足動物的內臟和體腔演化的意見促使HYMAN (1959, p. 230)認為觸手冠動物“構成了連接原口動 物與后口動物的模式”。在對觸手冠動物幼蟲發育的研究和重新解釋的基礎上,NIELSEN (1995, p. 6)同樣反對觸手冠動物的單系性,並將苔蘚動物歸為原口而將腕足動物和帚蟲歸為後口。在他的看法里,很多三個類群中的共有特徵,例如觸手冠,不是同源派生特徵,而是非同源的相似。然而,動物學家的普遍傾向於將所有觸手冠動物視為後口動物。(ZIMMER, 1964; BRUSCA & BRUSCA, 1990; SCHRAM, 1991; MEGLITSCH & S CHRAM , 1991; E ERNISSE , A LBERT , & ANDERSON, 1992)
觸手冠
以上這些基於經典生物學數據的相互矛盾的觀點,其爭論或許可以被分子證據所解決。特別是通過核糖體RNA的應用,來確定後生動物親緣關係的分子系統發生學(CONWAY MORRIS, 1993)。到目前為止,通過比較18S rRNAs序列,用
舌形貝代表腕足動物,得出一致的結論,表明腕足動物屬於原口動物(FIELD & others, 1988; GHISELIN, 1988; PATTERSON, 1989; LAKE, 1990; ADOUTTE & PHILLIPE, 1993),雖然其在原口動物內部的分 類位置仍不確定。

觸手冠
然而,觸手冠動物更詳盡的
基因學研究,開始提出嚴峻的門內分類學問題。一個對有鉸類腕足和無鉸類腕足,以及帚蟲、苔蘚動物的序列分析表明有鉸類腕足和帚蟲是姐妹類群;另外,無鉸類腕足、
軟體動物和環節動物多毛類共同構成一個與苔蘚動物為姐妹類群的分支(HALANYCH & others, 1995)。為了反駁這種腕足動物雙起源的見解,一個對代表所有現生腕足超科中超過三十個種的更詳盡的18S rRNA研究,顯示它們是單起源的(COHEN & GAWTHROP, 1996)。儘管如此,COHEN和GAWTHROP將帚蟲確定地歸於腕足動物的系統樹中,作為所有無鉸類腕足和單獨髑髏貝類腕足的姊妹類群。他們的結果微弱地支持將軟體動物(有可能包括環節動物)作為所有腕足動物和帚蟲的姊妹類群。但是沒能表示苔蘚動物的親緣關係。
通過以上確證的研究,腕足動物被認為構成一個種系分支,應該被識別為一個
動物門。我們相信,最終
分類學的結果將使該門容納其他觸手冠動物,例如帚蟲。同時,它還容納了三個亞門和八個綱,診斷和討論它們適合的分類位置見系統分類學。